 |


Морфогенетические зародыши
|
|
|
|
Морфогенез не происходит в вакууме. Он может начаться только с уже организованной системы, которая служит морфогенетическим зародышем. В процессе морфогенеза новая морфическая единица более высокого уровня возникает вокруг этого зародыша под влиянием специфического морфогенетического поля. Так как же это поле соединяется с морфогенетическим зародышем, чтобы началось развитие?
Ответ может состоять в том, что как соединение материальных систем с гравитационными полями зависит от их массы, а с электромагнитными полями — от их электрического заряда, так и соединение систем с морфогенетическими полями зависит от их формы. Таким образом, морфогенетический зародыш окружается морфогенетическим полем благодаря тому, что он имеет свою характерную форму.
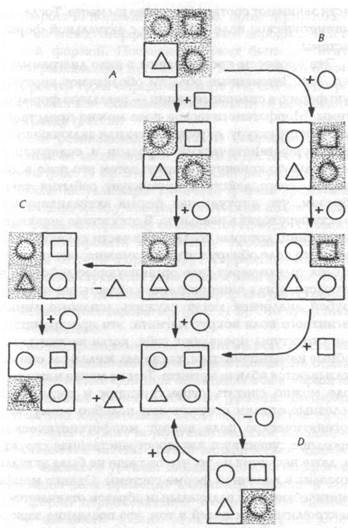
Рис. 11. Диаграмма, представляющая развитие системы из морфогенетического зародыша (треугольник) по нормальной хреоде (А). Показаны также альтернативный морфогенетический путь развития (В), регуляция (С) и регенерация (D). Затененные области представляют виртуальные формы внутри морфогенетического поля
Морфогенетический зародыш является частью системы, которая должна возникнуть. Поэтому часть морфогенетического поля системы соответствует этому зародышу. Однако остальная часть поля еще не занята или не заполнена; она содержит виртуальную форму законченной системы, которая становится реальной (актуальной) только тогда, когда все ее материальные части занимают соответствующие им места. Тогда морфогенетическое поле совпадает с актуальной формой системы.
Эти процессы представлены в виде диаграммы на рис. 11. Затененные области обозначают виртуальную форму, а сплошные линии — реальную форму системы. Морфогенетическое поле можно представить себе как структуру, окружающую или включающую в себя морфогенетический зародыш и содержащую виртуальную конечную форму; затем это поле в диапазоне своего действия организует события таким образом, что виртуальная форма актуализируется, то есть переходит в реальную. В отсутствие морфических единиц, которые составляют части конечной системы, это поле обнаружить невозможно; оно проявляет себя только через свое организующее действие на эти части, когда они попадают в область его влияния. Грубой аналогией могут служить «силовые линии» магнитного поля вокруг магнита; эти пространственные структуры проявляют себя, когда частицы, способные намагничиваться, такие как железные опилки, оказываются вблизи магнита. Тем не менее магнитное поле можно считать существующим и тогда, когда железные опилки отсутствуют; подобно этому, морфогенетическое поле вокруг морфогенетического зародыша существует как пространственная структура, даже несмотря на то, что она еще не была актуализирована в конечной форме системы. Однако морфогенетические поля радикальным образом отличаются от электромагнитных полей в том, что последние зависят от фактического (актуального) состояния системы — от распределения и движения заряженных частиц,— тогда как морфогенетические поля соответствуют потенциальному состоянию развивающейся системы и уже присутствуют раньше, чем система принимает свою конечную форму[104].
|
|
|
На рис. 11 показано несколько промежуточных состояний между морфогенетическим зародышем и конечной формой. Последняя может быть достигнута также различными путями {рис. 11 В), но если обычно выбирается один определенный путь, его можно рассматривать как «канализированный путь изменения», или хреоду (рис. 5).
Если развивающаяся система повреждена путем удаления ее части, она все же может достичь конечной формы (рис. 11 С). Это представляет регуляцию.
|
|
|
После актуализации конечной формы продолжающаяся связь между морфогенетическим полем и системой, форма которой ему соответствует, будет стремиться стабилизировать эту систему. Любые отклонения системы от этой формы будут корректироваться, так как система стремится вернуться к этой форме. А если часть системы удалить, конечная форма будет стремиться снова актуализироваться (рис. 11 D). Это представляет регенерацию.
Тип морфогенеза, показанный на рис. 11, существенно агрегативный: ранее существовавшие отдельно морфические единицы объединяются в морфическую единицу более высокого уровня. Возможен другой тип морфогенеза, когда морфическая единица, которая служит морфогенетическим зародышем, уже является частью другой морфической единицы более высокого уровня. Влияние нового морфогенетического поля приводит к трансформации, в которой форма первоначальной морфической единицы более высокого уровня заменяется формой новой морфической единицы. Большинство видов химического морфогенеза относится к агрегативному типу, тогда как биологический морфогенез предполагает обычно комбинацию трансформативного и агрегативного процессов. Примеры мы рассматриваем в следующих разделах.
Химический морфогенез
Агрегативные морфогенезы с нарастающей интенсивностью осуществляются в неорганических системах при понижении температуры: когда плазма охлаждается, субатомные частицы агрегируют в атомы; при более низкой температуре атомы агрегируют в молекулы; затем молекулы конденсируются в жидкости; и наконец, жидкости кристаллизуются.
В состоянии плазмы обнаженные атомные ядра можно рассматривать как морфогенетические зародыши атомов; они связаны с атомными морфогенетическими полями, которые содержат виртуальные орбитали электронов. В одном смысле эти орбитали не существуют, но в другом — они обладают реальностью, которая выявляется в остывающей плазме, когда они актуализируются путем захвата электронов.
Электроны, которые были захвачены атомными орбиталями, могут быть смещены оттуда воздействием внешней энергии или путем введения виртуальной орбитали с более низкой потенциальной энергией. В последнем случае они теряют дискретный квант энергии, который излучается как фотон. В атомах со многими электронами каждая орбиталь может содержать только два электрона (с противоположными спинами); таким образом, в остывающей плазме электронами заполняются сначала виртуальные орбитали с наиболее низкой энергией, затем орбитали с ближайшими к ним низшими энергиями и так далее, до тех пор пока не реализуется (актуализируется) полная атомная форма вокруг морфогенетического зародыша ядра.
|
|
|
Атомы, в свою очередь, являются морфогенетическими зародышами молекул, а малые молекулы — зародышами больших молекул. Химические реакции включают либо агрегацию атомов и молекул в большие молекулы — например, образование полимеров,— либо расщепление больших молекул на меньшие или на атомы и ионы, которые затем могут агрегировать с другими атомами или ионами, например в процессе горения: под действием внешней энергии молекулы распадаются на атомы и ионы, которые затем соединяются с кислородом с образованием малых, простых молекул, таких как Н2О и СО2. Эти химические изменения включают актуализацию виртуальных форм, связанных с атомами или молекулами, которые действуют как морфогенетические зародыши.
Мысль о том, что молекулы имеют виртуальные формы ранее, чем они актуализируются, особенно ясно иллюстрируется тем хорошо известным фактом, что совершенно новые соединения сначала могут быть «спроектированы» на основе эмпирически определенных принципов химического взаимодействия, а затем уже фактически синтезированы химиками-органиками. В этих лабораторных синтезах одна стадия следует за другой; на каждой стадии определенная молекулярная форма служит морфогенетическим зародышем для следующей виртуальной формы, которая должна быть синтезирована, и в конечном итоге образуется форма совершенно новой молекулы.
Если представление о химических реакциях как о морфогенетических процессах кажется несколько искусственным, то следует вспомнить о том, что действие катализаторов, как неорганических, так и органических, в значительной степени зависит от их формы. Например, ферменты, специфические катализаторы множества биохимических реакций, имеют поверхности, канавки, выемки или впадины, которым соответствуют реагирующие молекулы со специфичностью, часто сравниваемой с тем, как ключ соответствует замку. Каталитический эффект ферментов в большой степени зависит от того способа, которым они удерживают реагирующие молекулы в положениях, нужных для того, чтобы реакция произошла. (В свободном растворе случайные столкновения молекул происходят во всех возможных ориентациях, большинство из которых непригодны для реакции.)
|
|
|
Детали химического морфогенеза неясны отчасти из-за их большой скорости, отчасти потому, что промежуточные формы могут быть весьма нестабильны, а также потому, что конечные изменения состоят из вероятностных квантовых скачков электронов между орбиталями, которые образуют химические связи. Виртуальная форма молекулы, которая должна возникнуть, запечатлена в морфогенетическом поле, связанном с атомным или молекулярным морфогенетическим зародышем; когда другой атом или молекула приближаются в подходящей ориентации, форма образующейся молекулы актуализируется посредством квантовых скачков электронов на орбитали, которые ранее существовали лишь как виртуальные формы; в то же время при этом освобождается энергия, обычно в виде теплового движения. Роль морфогенетического поля в этом процессе является, так сказать, энергетически пассивной, но морфологически активной; оно создает виртуальные структуры, которые затем актуализируются, по мере того как морфические единицы низшего уровня «защелкиваются» («slot» or «snap») в эти структуры, освобождая при этом энергию.
Любой атом или молекула данного типа могут участвовать во многих видах химических реакций и поэтому являются потенциальными зародышами множества различных морфогенетических полей. Эти поля можно представить как возможности, «парящие» вокруг них. Однако атом или молекула могут не выполнить свою роль зародыша определенного морфогенетического поля до тех пор, пока к нему или к ней не приблизится подходящий атом или молекула, возможно, благодаря их специфическим электромагнитным или иным воздействиям.
Морфогенез кристаллов отличается от такового атомов и молекул тем, что определенный рисунок атомной или молекулярной организации повторяется неограниченно. Морфогенетический зародыш создается самим этим рисунком. Хорошо известно, что введение «семян» или «ядер» кристалла соответствующего типа значительно ускоряет кристаллизацию переохлажденных жидкостей или перенасыщенных растворов. В отсутствие таких семян или ядер морфогенетические зародыши кристаллов появляются только тогда, когда атомы или молекулы случайно занимают подходящие положения относительно друг друга вследствие теплового движения. Когда зародыш присутствует, виртуальные формы копий структуры решетки, задаваемой этим морфогенетическим полем, распространяются в разные стороны от поверхностей растущего кристалла. Подходящие свободные атомы или молекулы, которые приближаются к этим поверхностям, захватываются ими и «защелкиваются» в нужное положение; опять-таки при этом выделяется тепловая энергия.
|
|
|
Осеменение, или нуклеация, переохлажденных жидкостей может также проводиться, хотя и менее эффективно, путем введения малых фрагментов чужеродных веществ: например, химики часто скребут по краям пробирок, чтобы засеять растворы кусочками стекла. Эти кусочки служат поверхностями, позволяющими легче находить наиболее подходящие относительные положения тем атомам или молекулам, которые образуют истинный морфогенетический зародыш кристалла. По своим морфогенетическим эффектам такие семена напоминают катализаторы химических реакций.
Все виды химического морфогенеза, рассмотренные до сих пор, несомненно, агрегативны. В неживых  системах явления трансформации гораздо менее распространены. Например, большинство кристаллов не способны превращаться в другие кристаллические формы; они могут расплавиться или раствориться, и тогда их составляющие могут принять участие в других процессах кристаллизации; но это есть разрушение (дезагрегация), за которым следуют другие виды агрегации. Подобным же образом химические реакции включают дезагрегативные и агрегативные изменения. Однако существуют важные примеры молекулярной трансформации, такие как складывание белков, и обратимые изменения формы, которые происходят, когда некоторые ферменты связываются с молекулами, реакции которых они катализируют[105].
системах явления трансформации гораздо менее распространены. Например, большинство кристаллов не способны превращаться в другие кристаллические формы; они могут расплавиться или раствориться, и тогда их составляющие могут принять участие в других процессах кристаллизации; но это есть разрушение (дезагрегация), за которым следуют другие виды агрегации. Подобным же образом химические реакции включают дезагрегативные и агрегативные изменения. Однако существуют важные примеры молекулярной трансформации, такие как складывание белков, и обратимые изменения формы, которые происходят, когда некоторые ферменты связываются с молекулами, реакции которых они катализируют[105].
Тот факт, что белки складываются гораздо быстрее, чем можно было бы ожидать, если бы они «находили» свою конечную форму путем «случайного поиска», указывает на то, что их складывание происходит по определенным направлениям или по ограниченному числу направлений (раздел 3.3). Эти «канализированные пути изменения» можно рассматривать как хреоды. Чтобы процесс складывания мог начаться, согласно представлениям, излагавшимся выше в разделе 4.1, должен присутствовать морфогенетический зародыш и этот зародыш должен уже иметь характерную трехмерную структуру, которую он имеет в конечной форме белка. Предположение о существовании таких морфогенетических пусковых точек фактически уже было высказано в литературе по складыванию белков:
«Высокая скорость раскручивания предполагает необходимость того, чтобы процесс протекал по ограниченному числу направлений, даже когда статистика эта строго ограничена своего рода основными правилами стереохимии, которые содержатся в неявном виде в так называемом графике Рамачандрана. Становится необходимым постулировать в процессе складывания существование ограниченного числа допустимых инициирующих событий. Такие события, обычно называемые нуклеациями, вероятнее всего, происходят в тех частях полипептидной цепи, которые могут участвовать в конформационном равновесии между случайными и кооперативно стабилизированными структурами. Далее важно подчеркнуть, что аминокислотные последовательности полипептидных цепей, предназначенные для того, чтобы служить фабрикой белковых молекул, функционально осмыслены лишь тогда, когда они имеют трехмерную структуру, которая характерна для них в молекуле природного белка. Кажется разумным предположить, что части белковой цепи, которые могут служить как центры нуклеации для складывания, будут теми, которые могут "отклоняться" внутрь и наружу от конформации, которую они имеют в конечном белке, и что они образуют относительно жесткую структуру, стабилизированную набором кооперативных взаимодействий» [106].
Такие «центры нуклеации» будут действовать как морфогенетические зародыши через их связывание с морфогенетическим полем белка, которое затем будет канализировать процесс складывания по направлению к характерной конечной форме.
|
|
|
