 |


Вопрос 2. Непрерывные и синхронизированные структуры, способы их получения и назначения.
|
|
|
|
Существуют две принципиально разные системы выращивания микроорганизмов в жидкой среде. В одном случае после инокуляции среды не происходит ни добавления в нее, ни удаления каких-либо компонентов, кроме газовой фазы. Такая закрытая система культивирования носит название периодической и может поддерживать размножение клеток только в течение ограниченного времени, на протяжении которого меняется состав исходной среды и окружающие условия.
Непрерывное (проточное) культивирование в отличие от периодического характеризуется постоянной подачей питательной среды со скоростью, равной скорости удаления культуры. При этом объем культуры в ферментере во времени не меняется. Одно из основных условий непрерывного культивирования — хорошее перемешивание культуры в ферментере. Система непрерывного культивирования может быть реализована по принципу турбидо-стата или хемостата. Турбидостат — наиболее простой режим проточного культивирования, концентрация клеток в нем выбирается исследователем, а поступление питательных компонентов автоматически реализуется в соответствии с плотностью популяции. Меняя скорость подачи питательной среды («скорость разбавления»), экспериментатор может получать разные значения скорости роста популяции — от близких к нулю до максимальной, таким образом воспроизводя разные состояния культуры от стационарной фазы до стадии.экспоненциального роста.
Для непрерывного культивирования микроорганизмов может быть использован слегка модифицированный ферментер, применяемый при периодическом культивировании. В первом случае требуется система двойного насоса (для добавления свежей среды и удаления культуральной жидкости), используемая вместе с регулятором уровня среды; при этом культуральная жидкость удаляется через отверстия для отбора проб. Такие насосы необходимы при проточном культивировании в больших емкостях. В малых ферментерах удаление жидкости может происходить
|
|
|

через боковую отводную трубку, расположенную на уровне, позволяющем поддерживать определенный объем культуры. Малые ферментеры с боковым ОТВОДОМ ЖИДКОСТИ можно изготовить из стандартных стеклянных или стальных сосудов.
типичная кривая роста простой гомогенной периодической культуры бактерий. Рост проходит через лаг-фазу, в течение которой число клеток не увеличивается. Затем начинается фаза роста, которая обычно характеризуется экспоненциальным увеличением количества клеток и подчиняется уравнениям (10.1) — (10.4). В конечном счете изменения химического и физического состава среды приводят к переходу культуры в стационарную фазу, в которой увеличения количества клеток не происходит, но клетки еще нуждаются в источниках энергии для поддержания своей жизнедеятельности. Поскольку в периодической культуре питательные вещества ограниченны, наступает фаза отмирания (автолиза), которая часто характеризуется экспоненциальным уменьшением количества жизнеспособных клеток.
Лаг-фазу можно вызвать путем быстрого изменения состава среды культивирования. В свежей среде длительность лаг-фазы зависит от количества и возраста посевного материала, а также от изменений состава и концентрации питательных компонентов при внесении инокулята. Внесение небольшого инокулята в большой объем свежей среды может привести к диффузии из клеток витаминов, кофакторов и ионов, которые необходимы для поддержания активности многих внутриклеточных ферментов. Если клетки инокулируют из богатой среды в минимальную, то продолжительность лаг-фазы очень сильно зависит от объема инокулята, поскольку он содержит микроэлементы, попадающие при инокуляции в ростовую среду.
|
|
|
Длительность лаг-фазы зависит и от возраста инокулята. Это связано с тем, что в клетках накапливаются токсичные вещества, но в них недостаточно питательных веществ, необходимых для первоначального роста. Таким образом, наблюдается как положительный, так и отрицательный эффект возраста инокулята на продолжительность лаг-фазы в свежей среде. Как правило, при переносе клеток из бедной среды в богатую с увеличением возраста инокулята лаг-фаза удлиняется. На кривой зависимости продолжительности лаг-фазы от возраста инокулята при пересеве культуры с богатой среды на более бедную можно обнаружить определенный «минимальный» участок, отражающий обмен между питательными веществами и внутриклеточными токсичными соединениями.
Наконец, изменения состава питательных компонентов, а также их концентраций при переносе инокулята на свежую среду могут действовать как триггер при регуляции активности ферментов и морфологической диф-ференцировке клеток, например при прорастании спор. Если клетки переносят с бедной среды на богатую, то питательные компоненты и время расходуются на повышение активности ферментов, необходимых для осуществления метаболизма в целом. Если же клетки переносят с богатой среды на среду с более низким уровнем питательных веществ, то они способны немедленно, хотя и с низкой скоростью, вступить в экспоненциальную фазу роста.
Возможность постоянного экспоненциального роста в периодической культуре, хотя и в течение ограниченного времени, свидетельствует о том, что его скорость фактически не зависит от изменений концентрации субстратов в довольно широких пределах. В этом случае можно говорить о сбалансированном росте культуры (см. выше), который описывается только величиной ц. Со временем рост культуры начинает отклоняться от экспоненциального, и его уже нельзя описывать только с помощью одной величины ц, даже если можно рассчитать эту величину для случая лимитирования субстратом [уравнение (10.5)]. Моно [44] следующим образом описал соотношение между концентрацией субстрата и ростом бактерий в простой системе, находящейся в экспоненциальной фазе:
|
|
|

где ц — удельная скорость роста, |imax— максимальное значение (л, полученное при S^>Ks> Ks — константа насыщения, эквивалентная константе Михаэлиса — Ментен, a S — концентрация субстрата во время экспоненциальной фазы роста.
Для описания зависимости роста культур от концентрации субстрата существует несколько альтернативных моделей [44]. Уравнение (10.5) можно использовать робно понятие экономического коэффициента обсуждается ниже в этой же главе (разд. 10.5). Чтобы предсказать время достижения максимальной плотности биомассы в периодической культуре, необходимо сделать ряд упрощающих допущений [2, 4].
В периодической культуре максимальную биомассу определяют из экспериментальных данных, исходя из отношения прироста массы клеток к соответствующему снижению концентрации субстрата: ) 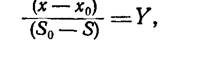
где х и S — соответственно концентрации клеток и субстрата в момент времени /, Хо и S0 — концентрации клеток и субстрата в более раннее время /0, а У — коэффициент общего выхода (экономический коэффициент). Уравнение точно описывает соотношение между концентрациями клеток и субстрата во время экспоненциальной фазы роста. Если экспоненциальный рост продолжается с постоянной скоростью вплоть до достижения стационарной фазы, а субстрат полностью исчерпывается во время экспоненциального роста, то с помощью уравнения (10.6) можно рассчитать максимальное число клеток или их концентрацию. Этот расчет верен при условии, что экономический коэффициент остается на постоянном уровне в течение всего цикла роста и потреблением субстрата в течение лаг-фазы можно пренебречь. На самом деле экономический коэффициент остается постоянным лишь при условии постоянного экспоненциального роста с одной удельной скоростью. Отсюда следует, что предсказанная с помощью уравнения (10.6) величина максимальной биомассы явно завышена.
|
|
|
