 |


Нервная ткань. Нейроциты и нейроглия. Нервные волокна
|
|
|
|
ОБЩАЯ МОРФОФУНКЦИОНАЛЬНАЯ ХАРАКТЕРИСТИКА. Нервная ткань относится к специализированным тканям. В филогенезе она возникла позже тканей общего значения в связи с усложнением строения и поведения животных. Выполняет важнейшую функцию — функцию реактивности. Эта функция основана на способности нейроцитов воспринимать раздражения, формировать нервные импульсы и вызывать ответные реакции. Из нервной ткани образуется нервная система, которая выполняет функцию хранения и переработки информации, регулирует и интегрирует все системы организма, осуществляет связь его с внешней средой.
Источником развития нервной ткани является нервная пластинка — часть эктодермы (нейроэктодерма). Из нее образуются два основных зачатка, дающие нервную ткань: нервная трубка и нервный гребень (ганглиоз-ные пластинки). Тканевыми элементами нервной ткани являются два вида клеток: нейроциты, или нейроны, и клетки нейроглии. Нейроциты являются ведущими клетками нервной ткани, ответственными за выполнение всех ее функций. Нейроглия но отношению к нейроцитам выполняет вспомогательные функции: трофическую, барьерно-защитную, опорную, регулятор-ную и др. Нейроциты нервной ткани взрослого организма не имеют недифференцированных предшественников, поскольку в ходе эмбриогенеза все первоначально способные к делению клетки подвергаются необратимой терминальной дифференцировке, нейроны взрослого человека являются высокоспециализированными клетками и не могут делиться. Поэтому регенерация нейроцитов на клеточном уровне невозможна. Она осуществляется исключительно на внутриклеточном уровне и в силу большой функциональной нагрузки на нейроциты протекает весьма интенсивно. Вариантом внутриклеточной регенерации является регенерация нервных волокон после их повреждения. Единственным исключением из этого правила являются обонятельные нейроны, способные к пролиферации. В остальном же на клеточном уровне регенерирует только нейроглия, имеющая камбий. Нервная ткань хорошо кровоснабжается, кровеносные сосуды лежат в прослойках РВНСТ.
|
|
|
ИСТОЧНИКИ РАЗВИТИЯ И ГИСТОГЕНЕЗ НЕРВНОЙ ТКАНИ. Источником развития нервной ткани является нейроэктодерма — часть эктодермы наружного зародышевого листка, имеющая вид дорзального утолщения, лежащего над хордой. Она называется нервной пластинкой. Детерминация клеток нервной пластинки происходит под влиянием индукции, исходящей от хордо-мезодермалыюго участка в ходе второй фазы гастру-ляции. В результате нейруляции, которая протекает на 18—21-е сутки эмбриогенеза, материал нервной пластинки разделяется на три составные части: нервную трубку, ганглиозные пластинки (нервный гребень) и нейроген-ные плакоды. В результате центрального изгиба нервной пластинки вначале образуется нервный желобок с утолщенными и приподнятыми краями. В процессе нейруляции изгиб желобка нарастает, его края сближаются и, наконец, срастаются. Формируется нервная трубка (22-е сутки эмбриогенеза). Она смещается под кожную эктодерму, полностью отделяясь от нее. При образовании нервной трубки часть клеток нервной пластинки формируют ганглиозные пластинки (нервный гребень). Одновременно по краям от нервной трубки в краниальном отделе зародыша формируются утолщения эктодермы, которые называются нейрогенными плакодами.
Нервная трубка и нервный гребень — основные зачатки, из которых развивается нервная ткань. Из нервной трубки развиваются нейроциты и макроглия центральной и периферической нервной системы. Из клеток нервного гребня образуются нейроциты и макроглия спинномозговых и вегетативных нервных узлов, узлов некоторых черепномозговых нервов, мозговое вещество надпочечников, меланоциты и клетки диффузной эндокринной системы. Из нейрогенных плакод образуется сенсорный и покровный эпителий органов вкуса, слуха и равновесия, а также нейроциты ганглиев V, VII, IX и X пар черепномозговых нервов.
|
|
|
Нервная трубка состоит из 5 слоев: 1) внутренней пограничной мембраны; 2) эпендимного; 3) плащевого (мантийного) слоев; 4) краевой вуали и 5) наружной пограничной мембраны. Эпендимный слой состоит из матричных (вентрикулярных) клеток, которые интенсивно делятся митозом, в результате чего число клеток нарастает. Заверпгившие пролифератив-ные процессы клетки эпендимного слоя переселяются в плащевой слой, однако при этом часть клеток остается на месте и служит для образования эпен-димной глии. Из клеток, переселившихся в мантийный слой, в результате детерминации образуются две линии клеточной дифференцировки: нейроген-ная и глиогенная (спонгиогенная). Нейрогенная линия дает нейроциты, из глиогенной образуются все виды макроглии, за исключением эпендимной глии. Краевая вуаль образована отросками клеток двух предыдущих слоев.
Стадии развития нейроцитов следующие: медуллобласт эпендимного слоя —> нейробласт —> пронейроцит —> нейроцит. Превращение медуллоблас-та в нейробласт происходит под влиянием нейромодуллина GAP-43. Пейромодуллин является белком, специфическим для аксона. Появление этого белка в клетках свидетельствует о начале дифференцировки. При этом пейробласт теряет способность к делению. На ранних этапах дифференцировки в нейробластах образуется несколько коротких отростков, причем потенциально каждый из них способен превратиться как в аксон, так и в дендрит. Накопление в отростке нейромодуллнна GAP-43 приводит к превращению отростка в аксон.
Нейробласт характеризуется наличием одного отростка (аксона) и синтезом нейрофибрилл. В его цитоплазме содержатся развитая гранулярная Э11С, комплекс Гольджи, многочисленные митохондрии. Нейробласты активно и строго целенаправленно мигрируют. Пронейроцит, или молодой нейроцит, быстро увеличивается в размерах, в нем появляются дендриты. большое число органелл, формируются синапсы с другими нейроцитами. Стадия зрелого нейроцита самая длительная. В эту стадию нейроцит приобретает дефинитивное строение и форму. Увеличивается число межней-рональных синапсов.
|
|
|
Целенаправленная миграция аксонов формирующихся нервных клеток обеспечивается в основном двумя механизмами. 1) Наличием особой разновидности энепдимной глин — таницитов, или радиальной глии. Таници-ты имеют радиальные отростки, вдоль которых но спирали мигрируют нейробласты. 2) Наличием хемотропизма отростков. Представления о нем были выдвинуты С. Рамоном-и-Кахалем. Теория хемотропизма получила полное подтверждение и называется теорией "меченых путей" или "верстовых столбов". Эти "столбы" представляют собой специфические молекулярные метки, образованные молекулами клеточной адгезии (МКА: лами-нин, фибронектип, коллаген и др.), которые последовательно "узнает" мигрирующий аксон-пионер, также содержащий в цитолемме МКА. Одной из МКА аксонов нейробласта является нейромодуллин GAP-43. За аксоном-пионером мигрируют аксоны других нейробластов, что ведет к развитию нервных трактов и нервов. Рост аксонов прекращается после достижения ими органов-мишеней.
Клетки нервных гребней мигрируют в несколько потоков в вентральном и латеральном направлении и, достигнув конечной точки миграции, под влиянием микроокружения дифференцируются в нейроциты, глиоци-ты ганглиев, либо в клетки мозгового вещества надпочечника, либо в ме-ланоциты.
Значительная часть нейроцитов в ходе гистогенеза нервной ткани погибает путем апоптоза (от 25 до 80%). Гибели подвергаются нейроциты, не установившие связи с органами-мишенями и не получившие от них специфических трофических факторов. Кроме того, гибнут и нейроны, установившие связь с органами-мишенями, но сформировавшие неправильные межнейронные связи. Показано, что в ходе гистогенеза нервной ткани первоначально образуется заведомо большее, чем необходимо, количество нейроцитов. Это создает определенную степень свободы для гистогенетичес-ких процессов. Затем лишние, с аномальными связями или не достигшие органа-мишени нейроциты подвергаются гибели.
|
|
|
При диффереицировке клеток глиогенной линии вначале образуются глиобласты. Из глиобластов образуется астроцитарная глия и олигоденд-роглия. Стадии развития глиоцитов такие: глиобласт —> проглиоцит —» гли-оцит (олигодендроглиоцит, эпендимоцит, астроцит). Из моноцитов крови, которые выселяются из сосудов, образуется микроглия. По некоторым представлениям, популяция клеток микроглии гетерогенна по происхождению. Одна ее часть образуется из моноцитов крови, тогда как другая имеет ней-роэктодермальное происхожден ие.
НЕЙРОЦИТЫ
КЛАССИФИКАЦИЯ. Существует несколько принципов классификации нейроцитов.
1. Морфологическая классификация (по числу отростков и форме пе-
рикарионов):
1.1. Униполярный нейроцит имеет один отросток аксон, который далее
ветвится на несколько отростков.
1.2. Биполярный нейроцит имеет два отростка: один из них аксон, второй — дендрит.
1.3. Псевдоуниполярный нейроцит. Это нейроцит, от тела которого отходит один отросток, который затем делится на два отростка (рис. 1.1 а). Некоторые авторы не выделяют униполярные нейроциты как самостоятельную структурную группу, считая их разновидностью биполярных нейронов.
1.4. Мультиполярные нейроциты. Имеют несколько отростков, один из которых аксон, а остальные дендриты. Этот вид нейронов — самый распространенный вид нейронов в организме взрослого человека (рис. 13.1 б-г). Униполярные нейроны встречаются только у низших животных. Иногда к униполярным нейроцитам относят так называемые амакриновые нейроциты сетчатки глаза и межклубочковые нейроциты обонятельной луковицы. Униполярными являются также нейробласты. Биполярные нейроциты находятся в сетчатке глаза, спиральном и вестибулярном ганглиях, а псевдоуниполярные находятся только в снинальных ганглиях. Как биполярные, так и псевдоуниполярные нейроциты по функции являются чувствительными либо (значительно реже) вставочными (например, вставочными являются биполярные нейроциты сетчатки глаза). По форме перикариона нейроциты делятся на звездчатые, пирамидальные, грушевидные, веретеновидные, паукообразные и др. (грушевидные и пирамидные нейроциты показаны на рис. 13.1 в, г).
2. Функциональная классификация. Учитывает выполняемые нейронами функции. Различают двигательные, чувствительные, ассоциативные и
нейросекреторные нейроциты.
2.1. Моторные, или двигательные (эфферентные, эффекторные), нейроны передают сигналы на рабочие органы (скелетные мышцы, железы, сосуды).
|
|
|

2.2. Чувствительные, или афферентные (сенсорные), нейроны. Их денд-риты заканчиваются чувствительными нервными окончаниями, воздействие на которые специфических раздражителей приводит к генерации нервного импульса, передаваемого далее по аксону либо на моторные, либо (значительно чаще) на ассоциативные
нейроциты.
2.3. Вставочные (ассоциативные нейроны, или интернейроны). Наиболее многочисленная (около 99,98% от всех нейроцитов) группа нейро-цитов, осуществляющих связь между нейронами.
2.4. Нейросекреторные нейроны. Это группа нейронов, специализированных на секреторной функции.
3. Медиаторная классификация. Нейромедиатор — это вещество химической природы, которое вырабатывается для передачи нервного импульса с одной нервной клетки па другую. Медиаторы могут иметь различную химическую природу, поэтому различают несколько типов нейронов:
3.1. Холинергические нейроны (медиатором является ацетилхолин).
3.2. Аминергические нейроны (медиаторами служат биогенные амины).
Они делятся на:
а) адренергические нейроны (медиатором является норадреналин);
б) серотонинергические нейроны имеют в качестве медиатора серо-
тонин
в) дофаминергические нейроны (медиатором является дофамин).
3.3. Пуринергические нейроциты передают нервные импульсы при помощи АТФ и других пуриновых оснований.
3.4. Пептидергические нейроны (в качестве медиаторов используются различные пептиды).
3.5. ГАМКергические нейроциты (медиатор — гаммааминомасляная
кислота, ГАМ К). Строение нейроцита. Нейроцит — это нервная клетка со всеми ее отростками и концевыми ветвлениями — нервными окончаниями. (Иногда студенты понимают под нейроцитом только перикарион, что в корне неправильно!). Отростки нервной клетки делятся на два вида: аксон (нейрит) и дендриты. По аксону нервный импульс идет от тела клетки к периферии, по дендритам возбуждение передается с периферии к телу клетки. Нейроциты могут иметь только один аксон и один или множество дендритов.
Ядро нейроцитов крупное, округлое, имеет одно крупное ядрышко (иногда может быть 2—3 ядрышка) (рис. 13.2). В ядре нейроцита преобладает эухроматин. У лиц женского пола около ядрышка выявляются тельца Барра, представляющие собой инактивированную Х-хромосому. Описанная структура ядра характерна для клетки с высокой белоксинте-зируемой функцией. Обычно нейроцит имеет одно ядро, но в нсйроцитах вегетативной нервной системы может быть более 10 ядер.
Цитоплазма нейроцита подразделяется на перикарион (часть цитоплазмы, окружающая ядро) и аксоплазму, или цитоплазму отростков. В пери-карионе при световой микроскопии основными красителями выделяется базофильная субстанция (тигроид, субстанция Ниссля). Она выявляется в теле нейрона, в дендритах, но отсутствует в аксоне и аксон-ном холмике — месте отхождения аксона от перикариона. В зависимости от функционального состояния нейроцита величина, расположение и количество глыбок базофильиого вещества могут изменяться. Оно может полностью исчезать, например, при регенерации нервных отростков. Явление исчезновения базофильиого вещества называется хроматолизом или тигролизом. В электронном микроскопе установле- / но, что базофильное вещество /Й — это сильно развитая грану- |w«лярная ЭПС, компоненты кото- щ рой лежат плотно и упорядо- Г', ченно, анастомозируя друг с \\ другом. Функция ее — биосин- * тез белка.
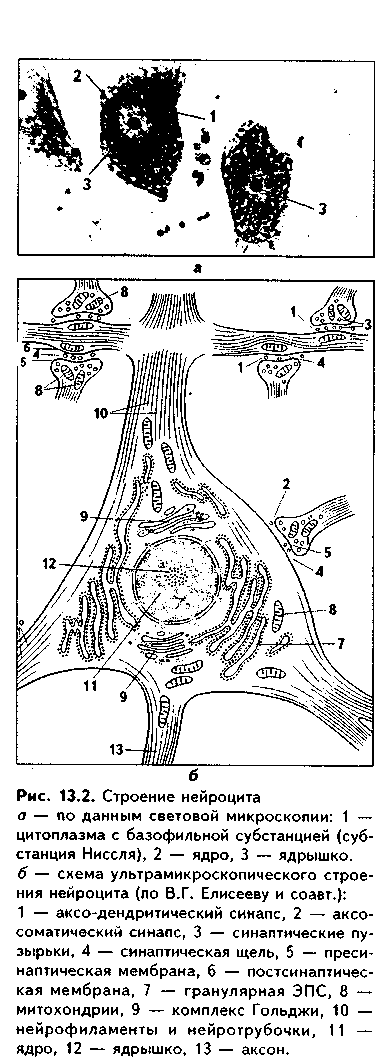
|
В цитоплазме перикариона при окраске азотнокислым серебром выявляются нейрофиб- _ риллы. Это нити толщиной от 0,5 до 3 мкм. Они идут в раз- ~ ных направлениях в перикари-оне и в отростках нейроцита и L, представляют собой компоненты цитоскелета, склеившиеся в пучки при фиксации материа- | ла (т.е. фибриллы по своей сути являются артефактом) (рис. 13.3). В электронном микроскопе показано, что ци-тоскелет нейроцитов представлен микротрубочками (нейртрубочками), микрофиламента-ми и промежуточными фила-ментами (нейрофиламентами). Микротрубочки и микрофила-менты имеют такое же строение, как и в других клетках. При этом нейротубулы при помощи специального белка кинезина связаны с органеллами незина связаны.
нейроцита и участвуют в аксональном токе.

|
Нейрофиламенты — это фио-риллярные структуры диаметром 6—10 нм, состоящие из лежащих по спирали белковых молекул. При помощи поперечных мостиков они связаны друг с другом и с нейротрубочками поперечными мостиками. Кроме перечисленных органелл в нейроците содержится большое число митохондрий, хорошо развиты комплекс Гольджи, гладкая ЭПС. Между ядром и дендритами лежат цснтриоли. В стареющих нейронах встречаются жировые и пигментные включения, в частности, липофусцин (неправильно называемый пигментом старения). Он представляет собой видоизмененные лизосомы и встречается даже в нейронах плодов. В некоторых нейроцитах (нейроциты голубого пятна, черной субстанции) находятся включения меланина. Лизосомальный аппарат нейроцитов выражен очень сильно, лизосомы имеют различные размеры, осуществляют разрушение стареющих компонентов цитоплазмы нейроцита (аутофагия), взамен которых образуются новые. Следовательно, лизосомы участвуют в постоянном обновлении компонентов цитоплазмы нейроцита (внутриклеточной регенерации).
Дендриты. Это отростки нейроцита, по которым нервный импульс передается к перикариону. Благодаря дендритам нейроцит получает информацию от других нейроцитов и от нервных окончаний. В области аксо-дендритических связей (синапсов) имеются дендритные выпячивания — так называемые дендритные шипики, в которых могут выявляться несколько цистерн, разделенных электронноплотным материалом. Цистерны и электронноплотный материал представляют собой шипиковый аппарат. Дендритные шипики очень лабильны, постоянно разрушаются и вновь образуются. Их количество существенно возрастает при усилении функциональной нагрузки на нейроцит и снижается при старении и падении функциональной нагрузки. Усиленное новообразование шиииков идет в первые месяцы жизни ребенка. В дендритах, особенно у места их отхождения от тела нейроцита, встречаются все виды органелл, количество которых падает по мере ветвления дендрита.
Аксон. Передает нервный импульс от тела нейроцита к другим нервным клеткам или на рабочий орган. Имеет размеры до 1,5 м и может содержать до 99% цитоплазмы. Начинается аксон от аксонного холмика — выпячивания перикариоиа, в котором находится комплекс Гольджи и отсутствует субстанция Ниссля. В аксонном холмике происходит генерация нервного импульса. В этой зоне цитолемма нейроцита обладает большим количеством ионных каналов, необходимых для деполяризации. В центре аксона проходят продольно ориентированные пучки нейрофиламентов, а более периферически находятся нейротубулы и микрофиламенты, другие органеллы: цистерны агранулярной ЭПС, элементы комплекса Гольджи, митохондрии.
Аксональный ток (аксоток). Основные синтетические процессы в нейроците идут в перикарионе. Здесь же сосредоточены основные органеллы. В отрсклках синтетические процессы идут медленнее и менее интенсивно. Поэтому вещества и органеллы поступают в отростки из перикариоиа. Установлено непрерывное движение нейроплазмы от тела клетки к термина-лям. Это движение называется аксотоком (термин распространяется как на движение веществ по аксону, так и по дендриту). Различают анте- роградный и ретроградный аксоток. Антероградный аксоток — это движение аксонлазмы от перикариоиа к терминальным ветвлениям. В свою очередь, антероградный аксоток подразделяется на медленный и быстрый аксоток. Медленный аксоток происходит со скоростью 1—5 мм в сутки. Посредством медленного аксотока транспортируются компоненты гиалоплазмы (аксоплазмы) с ферментами, а также компоненты цитос-келета. Быстрый аксоток протекает со скоростью от 50 до 2000 мм в сутки. Служит для транспорта большинства органелл и пузырьков медиаторов. Ретроградный аксоток — это аксоток от терминалей к перикариону. Имеет скорость до 200 мм в сутки. При помощи его к перикариону доставляются вещества, синтезируемые глией, из терминалей отростков удаляются различные вещества, транспортируются синаптические пузырьки, при помощи которых перикарион получает информацию о состоянии периферии. При помощи ретроградного аксотока могут транспортироваться стареющие органеллы, которые в дальнейшем подвергаются разрушению
лизосомами перикариоиа.
Механизм аксотока. В настоящее время считают, что структурную основу аксотока составляют нейротубулы, с которыми связаны сократимые белки ди-неин и кинезин. В связи с этим выделяют два основных механизма аксотока:
1. актин-миозиновый. На поверхности нейротубул обнаружены белки типа актина и миозина. Актин также входит в состав актиновых микро-филамент. Актиновые нити вступают в контакт с миозиновыми филамен-тами. К миозиновым филаментам прикрепляются пиноцитозные пузырьки. При помощи их совершается аксоток, они содержат вещества, доставляемые на периферию. В результате скольжения актиновых нитей вдоль миозиновых происходит транспорт пузырьков, происходит аксоток. Установлено, что вокруг нейротрубочек и нейрофиламентов находится менее вязкая зона аксоплазмы, что способствует транспорту везикул. В аксотоке играет роль также гладкая ЭПС, которая является источником образования транспортных пузырьков.
2. Тубулиново-кинезиновый (динеиновый) механизм аксотока заключается в следующем. Молекула кинезина одним концом прикрепляется к транспортируемой органелле или транспортному пузырьку, а другим совершает шаговые перемещения вдоль микротрубочки. Шаг перемещения кинезина составляет 8 нм. Сходным с кииезиновым является динеиновый
механизм аксотока.
Кроме указанных механизмов, в аксотоке играет роль глия, в первую очередь, олигодендроглия, клетки которой находятся в постоянных пульсирующих движениях. Это как бы массирует отростки нейроцитов и способствуют аксотоку. Функции аксотока:
1. В нейроне большинство веществ образуется в перикарионе, там же образуются и органеллы, с аксотоком они идут в отростки и обеспечивают их функции.
2. С ретроградным аксотоком в перикарион поступает информация с периферии, в том числе и в виде веществ, синтезируемых в глиальных клетках.
3. Аксоток играет важную роль в регенерации нервных волокон.
Роль аксотока в патологии. Патология аксотока. За счет аксотока могут транспортироваться не только метаболиты и органеллы, но и вирусы бешенства, герпеса, полиомиелита. Это способствует достаточно быстрому распространению данных микроорганизмов и поражению нейроцитов. Аксоток может нарушаться при недостатке витамина В, (болезнь бери-бери), при сахарном диабете, при подагре. Это ведет к дегенеративным изменениям нервных отростков и сопровождается потерей или понижением чувствительности, движений.
Плазмолемма (аксолемма) — это плазматическая мембрана нейроци-та. Она имеет такое же строение, как в других клетках, но в функциональном отношении имеет особенности: обладает повышенной способностью пропускать ионы, которые перемещаются за счет работы энергозависимого калий-натриевого насоса (энергозависимые ионные каналы). Он создает внутри клетки более высокую концентрацию ионов калия и более низкую концентрацию натрия по сравнению с внеклеточной средой. В покое происходит постепенная утечка калия во внеклеточную среду, что создает потенциал покоя в -70 мВ. При раздражении мембрана быстро пропускает натрий внутрь клетки, а калий — наружу. Возникает потенциал действия, или нервный импульс. Его генерация происходит в области аксонного холмика. Более подробно механизмы генерации нервных импульсов рассмотрены в курсе физиологии.
НЕЙРОГЛИЯ
В процессе развития нервной ткани из материала нейрального зачатка (нервной трубки, нервного гребня) наряду с нервными клетками образуются и глиоциты — вспомогательные клетки нервной ткани. Количество глиальных клеток как минимум в 10 раз превышает число нейроцитов. Термин "глия" (в переводе с греческого означает "клей") предложил известный немецкий патолог Р. Вирхов, который считал, что при помощи глии происходит склеивание нейроцитов в единое целое и заполнение промежутков между нейроцитами и нервными волокнами (по первоначальному представлению Р. Вирхова, глия является неклеточным материалом). Лишь позже была доказана клеточная природа глии. Глиальные клетки, в отличие от нейроцитов, способны к делению. Эта способность особенно возрастает при повреждении мозга, что ведет к формированию глиальных рубцов. Кроме того, из-за способности к пролиферации глиоциты могут формировать доброкачественные и злокачественные опухоли мозга (опухоли, происходящие из нейроцитов, точнее, из медуллобластов, возможны только в раннем постнатальном периоде).
Клетки глии выполняют трофическую, опорную, разграничительную, защитную, секреторную функции, участвуют в проведении нервного импульса, поддерживают гомеостаз нервной ткани, участвуют в образовании гемото-энцефалического барьера. Как видно из таблицы, нейроглия делится на две разновидности: мак-роглию и микроглию, или глиальные макрофаги. В свою очередь, макроглия делится на эпендимоглию, астроглию, олигодендроглию. Некоторые ученые указывают на наличие четвертого вида макроглии — мультипотен-циальной, способной превращаться в другие виды макроглии. По другим взглядам, так называемая мультипотенциальная глия представляет собой камбий для макроглии.

|
Эпендимная глия. Выстилает центральный канал спинного мозга, полости желудочков головного мозга (рис. 13.4 б). Эта глия имеет вид однослойного эпителия (по Н.Г. Хлопину, эпителий эпендимоглиального типа). На поверхности, обращенной в сторону канала, на глиальных клетках есть реснички. От базалыюй части клетки отходят отростки, которые идут через всю толщину спинного или головного мозга и соединяются друг с другом на наружной поверхности, участвуя в образовании наружной гли-альной пограничной мембраны. Боковыми сторонами эпенимоциты связаны друг с другом при помощи межклеточных контактов.
В области сосудистых сплетений, секретирующих спинномозговую жидкость, находится разновидность эпендимоглии, называемая хороидной эпендимоглией. Ее клетки имеют кубическую форму и покрывают выпячивания мягкой мозговой оболочки, вдающиеся в просвет желудочков го-ловкого мозга. Апикальные поверхности хороидных эпендимоцитов имеют многочисленные микроворсинки, базальные формируют многочисленные ножки, переплетающиеся и образующие своеобразный базальный лабиринт. Боковыми поверхностями клетки тесно связаны друг с другом.
Танициты, находящиеся в стенках 3-го желудочка, воронкового кармана и срединного возвышения, также относятся к эпендимоглии. Танициты имеют кубическую или призматическую форму. На апикальной поверхности несут микроворсинки и отдельные реснички. От базальной поверхности клеток отходит отросток, идущий к капилляру и образующий на нем пластинчатое расширение. Радиальные глиоциты как разновидность эпендимоглии описаны выше. Все глиоциты лежат па базальной мембране.
Функции эпендимоглии: опорная, защитная, секреторная (секреция церебральной жидкости), разграничительная, защитная, трофическая. Эиендимоглия образует нейро-ликворный и гемато-ликворный барьеры (соответственно барьеры между нейроцитами и ликвором, кровью и ликво-ром). Танициты осуществляют транспорт веществ из ликвора в кровеносные сосуды, осуществляют тем самым связь между этими двумя жидкимисистемами.

|
Астроцитная глия. Составляет опорную структуру головного и спинного мозга. Маркером астроглии является глиальный фибриллярный кислый белок (ГФКБ), из которого построены промежуточные филаменты. Есть две разновидности астроглии: плазматическая и волокнистая. В сером веществе преобладает плазматическая астроглии, а в белом — волокнистая. Плазматическая глия имеет короткие и толстые отростки (рис. 13.4 а), богатые цитоплазмой различными органеллами, включениями гликогена и с невысоким содержанием промежуточных филаментов. Волокнистая астроглия имеет тонкие длинные отростки, в которых содержится большое количество фибриллярного аппарата. За счет отростков глиоцитов создаются глиаль-ные опорные и разграничительные структуры (мембраны) в белом веществе. При помощи отростков астроциты контактируют не только друг с другом, но также с клетками олигодендроглии и эпендимоглии. Плазматическая астроглия создает глиальные пограничные мембраны вокруг сосудов и участвует в образовании гематоэнцефалического барьера (ГЭБ).

Функции астроглии: опорная; барьерно-защитная (участие в образовании ГЭБ; астроциты имеют выраженную способность к фагоцитозу, переработке и представлению антигенов, выработке медиаторов иммунных реакций); разграничительная; транспортная (участие в аксотоке); трофическая; регуляторная и метаболическая (астроциты способны захватывать медиатор из синаитической щели и передавать его нейрону, участвуют в метаболизме медиаторов); пластическая (при повреждении мозга формируют глиальный рубец).
Олигодендроглия. Эта разновидность глии имеет небольшое число тонких отростков (в переводе с греческого термин "олигодендроглия" означает "глия с малым количеством отростков"). Тела клеток имеют неболь-раной. Некоторые исследователи отмечают также наличие в ЦНС очень тонких "голых" нервных волокон, полностью лишенных глиальной оболочки. Они чаще встречаются в период эмбриогенеза. Скорость проведения нервного импульса по безмиелиновым нервным волокнам невысока — 1—5 м/сек. Находится этот тин нервных волокон в основном в вегетативной нервной системе (пост-ганглионарные волокна).

|
Миелиновые нервные волокна также состоят из нервного отростка — осевого цилиндра—и леммоцитов. Отросток не просто лежит в углублении на поверхности леммоцита, а окружен слоистой оболочкой, образованной при накручивании мезаксона вокруг отростка нейроцита. Эта оболочка называется миелиновой оболочкой. Миелиновая оболочка состоит из внутреннего, собственно ми-елинового слоя, образованного многочисленными (до 200— 300) кольцами дубликатуры цитолеммы леммоцита, и наружного слоя,в котором находятся ядра и цитоплазма леммоцита — неврилеммы. Миелиновая оболочка содержит большие количества липидов и поэтому интенсивно окрашивается осмиевой кислотой. В отдельных участках в витках мезаксона между двумя его слоями остаются небольшие участки цитоплазмы. Эти уча
стки не прокрашиваются осмиевой кислотой и выглядят в виде расположенных иод острым углом к осевому цилиндру светлых полосок (насечки миелина или насечки Шмидт-Лантермана).
По ходу мислинового волокна есть сужения — узловые перехваты Ранвье. Они представляют собой границы двух соседних леммоцитов. В местах узловых перехватов в каждом из контактирующих леммоцитов образуется кольцо из плотно лежащих микротрубочек, которые обеспечивают плотное прилегание леммоцитов к осевому цилиндру. Кроме того, участки соседних леммоцитов образуют многочисленные отростки с интердигитациями,
которыми взаимодействуют друг с другом. В области узлового перехвата осевой цилиндр расширяется, его цитолемма содержит повышенное количество натриевых каналов, отсутствующих в других участках волокна. Все расстояние между двумя соседними перехватами называется межузловым сегментом.

|
Проведение нервного импульса по миелиновым нервным волокнам происходит в зависимости от толщины волокна со скоростью от 10 до 120 м/сек. Такая скорость обеспечивается следующим. Миелиновая оболочка действует наподобие аккумулятора — способствует накоплению электрического заряда. В области перехвата миелина нет, и весь заряд скапливается здесь. При достижении некоторого уровня он "перепрыгивает" на соседний перехват и затем на другие. Такой путь передачи импульсов называется сальтаторным (от слова сальто — прыжок). Миелиновые волокна толще безмиелиновых, причем каждое волокно содержит только один осевой цилиндр. Этот тип волокон находится в соматической нервной системе, входит в состав преганглионарных волокон ВНС. Снаружи и миелиновые, и безмиелиновые нервные волокна окружены базалыюй мембраной.
ОБРАЗОВАНИЕ МИЕЛИНОВЫХ НЕРВНЫХ ВОЛОКОН (МИЕ-ЛИНОГЕНЕЗ). Образование миелиповых нервных волокон несколько различается в центральной и периферической нервной системе (ПНС) (рис. 13.8 а).

|
В ПНС при образовании миелинового нервного волокна осе- вой цилиндр вдавливается в поверхность леммоцита, складки которого окружают осевой цилиндр и образуют мезаксон. Это первая стадия миелиноге-неза — стадия глиоза нервного волокна. Во вторую стадию (стадию мие- линогенеза) мезаксон начинает расти за счет синтеза леммоцитом все но- вых участков илазмолеммы и накручивается вокруг осевого цилиндра. Образуются многочисленные витки миелина. При этом цитоплазма леммоцита сдвигается на периферию вместе с ядрами леммоцита, образуя неврилемму. Снаружи от миелинового волокна образуется базальная мембрана. Между слоями мезаксона в некоторых местах остаются участки цитоплазмы (насечки миелина).
При миелинизации нервных волокон в ЦНС погружения осевого цилиндра в цитоплазму леммоцита не происходит. Вместо этого олигоденд-роцит формирует тонкий плоский отросток. Этот отросток в форме языка охватывает осевой цилиндр, а затем, в силу образования все новых порций цитолеммы, растет и послойно накручивается вокруг осевого цилиндра, образуя витки миелина (рис. 13.8 б). Второе отличие от миелинизации в ПНС состоит в том, что один олигодендроцит может принимать участие в миелинизации многих (до 50) осевых цилиндров, образуя столько же отростков. Зоны узловых перехватов в ЦНС более широкие, в отличие от ПНС перекрытия их цитолеммой леммоцитов не происходит. Это обеспечивает более высокую скорость передачи нервного импульса.

|
И в ЦНС, и в ПНС происходит постоянный процесс разрушения старых фрагментов миелина с замещением их новыми — ремоделирование миелина. Фагоцитоз старых компонентов миелина осуществляется как глиальными клетками, так и макрофагами РВНСТ эндоневрия.
ПАТОЛОГИЯ МИЕЛИНА. Физиологическая роль миелина особенно хорошо видна при рассмотрении так называемых демиелинизирующих заболеваний (рассеянный склероз, боковой амиотрофический склероз, сирин-гомиелия и др.), т. е. заболеваний, которые связаны с нарушением образования миелина и его повышенной деструкцией. Это может быть вызвано изменением химического состава миелина под действием некоторых вирусов (вирус кори и др.), и последующей аутоимунной реакцией на измененный миелин. Демиелинизация может наступать и при многих других заболеваниях (сахарном диабете, интоксикациях и др.). При дифтерии в периферической нервной системе также имеет место демиелинизация нервных волокон, однако она связана с тем, что дифтерийный токсин, не воздействуя на предсуществующий миелин, блокирует его синтез шван-новскими клетками, т.е. ремоделирование миелина. В любом случае потеря миелина ведет к нарушению изоляции нервных волокон, замедлению проведения нервного импульса и появлению тяжелых симптомов у больного.
|
|
|
