 |


Глава 2. Дисфагия, боли в груди и гастроэзофагальный рефлюкс
|
|
|
|
Тимоти Т. Нострант
Дисфагия и боль в груди — общая клиническая проблема, проявления которой могут серьезно влиять на качество жизни и часто требуют быстрой оценки и диагностики. Целью данной главы является ознакомление читателя с нормальной физиологией пищевода для того, чтобы использовать эти сведения в качестве основы для оценки пациентов с дисфагией и болями в груди. Кроме того, в главе представлены клинические примеры патофизиологических корреляций между дисфункцией пищевода и диагностическими признаками патологии.
Физиология глотания
Пищевод — простой полый орган, всегда остающийся пустым, несмотря на прием пищи и рефлюкс. В процесс продвижения пищи до пищевода вовлекаются ротовая полость, глотка и гортань, при этом попадание пищи в воздухоносные пути невозможно (рис. 2-1). Стенки верхних отделов глотки имеют три группы констрикторных мышц — верхнюю, среднюю и нижнюю. Данные констрикторы совместно с мышцами гортани предназначены для поднятия гортани от входа в пищевод, при этом мышцы средней и нижней части глотки сокращаются в заднепереднем направлении. Эти мышцы имеют плотную иннервацию, аналогичную по степени плотности иннервации окологлазных мышц (отношение нервных волокон к мышечным составляет от 1: 2 до 1: 6), что обеспечивает исключительно хорошую регуляцию. Центральный контроль и рефлекторная активация нейронов определяет последовательность мышечных сокращений и акт глотания.
Глотание — нейромышечная реакция с произвольным и непроизвольным компонентом (рис. 2-2). В среднем человек глотает 600 раз в сутки (200 раз во время еды, 50 раз во время сна, 350 раз в остальное время), преимущественно бессознательно. Наличие жидкой или твердой пищи в ротовой полости очень важно для процесса глотания, поскольку трудно глотать при совершенно пустой ротовой полости. Процесс глотания можно разделить на две фазы: ротовую и глоточную. Ротовая фаза преимущественно произвольная. Полость рта закрывается спереди губами, по центру языка формируется пищевой комок, и далее происходит его проталкивание назад до твердого неба. При этом в процессе глотания язык обеспечивает до 80 % энергии, требуемой для транспорта пищевого комка до пищевода. С задней поверхности языка пищевым комком запускается рефлекторный ответ глотки и последующие движения являются преимущественно непроизвольными. Рефлек
|
|
|
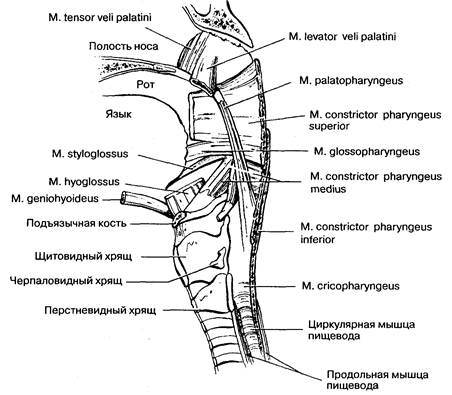
Рис. 2-1. Анатомия глотки. Глотка условно делится на три сегмента. (1) Носоглотка, которая не является частью пищеварительного тракта. М. levator veli palatini и m. tensor veli palatini закрывают глотку для предотвращения регургитации при глотании. (2) Верхняя часть глотки, где начинается пищеварительный и респираторный тракт. Ее основное предназначение — смещение глотки вверх и вперед. (3) Нижняя часть глотки наиболее важна в проталкивании пищевого комка и для движения его через верхний пищеводный сфинктер (rn. cricopharyngeus и m. constrictor pharyngeus inferior). (По: Yamada Т., Alpers D. H., Ovvyang С., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia: J. B. Lippincott, 1995: 160.)
торный ответ глотки состоит из 5 фаз и происходит в течение 1 с. Первая фаза — закрытие носоглотки в результате сокращения мягкого неба. Это предотвращает попадание комка в носовую полость и способствует его эффективному продвижению в нижние отделы глотки. Вторая фаза — передневерхнее смещение гортани, что предупреждает аспирацию. Чтобы и далее исключить попадание пищи в гортань, последняя закрывается надгортанником, истинными и ложными голосовыми связками, образуя форму закрытой чаши (третья фаза). Расслабление верхнего пищеводного сфинктера происходит после закрытия и смещения гортани и составляет четвертую фазу. Верхний сфинктер пищевода состоит из нижнего участка m. constrictor pharyngeus inferior и m. cricopharyngeus. Эти мышцы постоянно сокращены за счет непрерывной нервной стимуляции. Прекращение нейрональной активности вызывает снижение мышечного тонуса на 90 % вследствие прекращения нейрональной импульсации (10 % мышечного тонуса составляет базальный мышечный тонус). Последней фазой глотания является стимуляция сокращения глотки с последующим пассажем комка в открытый пищевод. Сфинктер прежде всего сокращается в переднезаднем направлении и образует щелеобразную конфигурацию
|
|
|
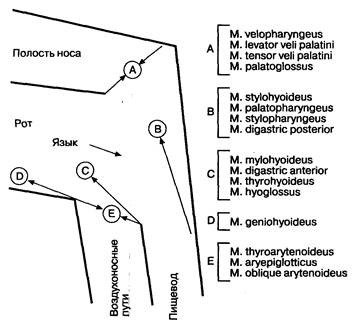
Рис. 2-2. Координация групп мышц. Мышцы группы А необходимы для защиты носоглотки от ретроградного пассажа пищевого комка. Мышцы группы В смещают гортань вверх от пути прохождения пищевого комка, в то время как группы мышц С, D и Е участвуют в смещении гортани вперед. Эти мышцы являются основными в процессе открытия В ПС. Проталкивание пищевого комка производится в основном языком, с небольшим участием мышц нижнего отдела глотки. (По: YamadaT., А1pers D. H., Owyang С., Роwell D. W., Silverstein F. Е., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia:.!. B. Lippincott, 1995:123.)
в форме "С" вокруг перстневидного хряща гортани. Тонус верхнего пищеводного сфинктера (ВПС) снижается во время сна и в этот период поддерживается только базальным мышечным тонусом, чтобы сфинктер находился в закрытом состоянии. Однако ВПС моментально реагирует на дыхание, фонацию, положение головы, растяжение, стимуляцию и напряжение, что защищает пищевод и воздухоносные пути.
Все стадии глотания зависят от характера пищевого комка. Для твердого пищевого комка требуется большее открытие ВПС и более сильное сокращение глотки. Для жидкой пищи — меньшее открытие ВПС и меньшая степень сокращения глотки. Кроме того, от размера и консистенции пищевого комка зависит продолжительность открытия ВПС. Для прохождения большого и твердого комка необходимо большее время открытия. Закрытие сфинктера совпадает с распространением сокращения по глотке, и тонус ВПС после глотка больше, чем его базальный тонус, что предотвращает ретроградный пассаж комка. Ретроградный выход газов (отрыжка) запускает расслабление ВПС растяжением тела пищевода, и продолжительность расслабления соответствует времени полного прохождения газов. Растяжение пищевода газом или баллоном вызывает расслабление ВПС, аналогичное естественному при отрыжке.
|
|
|
Пищевод представляет собой 20—22-сантиметровую мышечную трубку, стенки которой состоят из гладкой и поперечнополосатой мускулатуры: проксимальный отдел пищевода (5 %) состоит только из поперечнополосатой мускулатуры, в то время как средний его участок (35—40 %) содержит оба типа мышц; 50—60 % пищевода — дистальная часть, включая нижний пищеводный сфинктер (НПС), имеет только гладкую мускулатуру. В отличие от других отделов желудочно-кишечного тракта, пищевод не имеет серозной оболочки. Внешняя иннервация пищевода осуществляется через блуждающий нерв. Тела мотонейронов для поперечнополосатой мускулатуры пищевода находятся в n. ambiguus, в то время как тела нейронов, иннервирующих гладкую мускулатуру, находятся в задних двигательных ядрах n. vagus. Эти нейроны связаны с собственными нервными сплетениями пищевода, находящимися между продольным и циркулярным мышечными слоями. Данные сплетения более многочисленны в отделах пищевода с гладкой мускулатурой, чем с поперечнополосатой, но в целом их меньше в пищеводе, чем в других отделах желудочно-кишечного тракта. Нервные сплетения в подслизистом слое (сплетение Мейснера) в пищеводе почти не встречаются.
Перистальтика пищевода возникает сразу после того, как сокращение, начинающееся с глотки, пройдет через ВПС. Средняя скорость перистальтики — 2—4 см/с. Первичная перистальтика стимулируется глотанием, в то время как вторичная — запускается растяжением пищевода дистальнее области растяжения. Перистальтика в отсутствии внешней иннервации называется автономной, предполагая тем самым, что она может инициироваться только на интрамуральном уровне.
Другая особенность механизма перистальтики — способность угнетаться при глотании. Второй глоток полностью угнетает прохождение первого, если начинается до того, как первый глоток проходит часть пищевода, имеющую поперечнополосатую мускулатуру. Если первый глоток достигает дистальных отделов пищевода в момент второго глотка, то перистальтика первого глотка угнетается только частично. Короткие повторные глотательные движения вызывают полное угнетение пищевода с расслаблением НПС. Нормальная перистальтика возможна лишь при медленных глотках и полном освобождении пищевода от предыдущего пищевого комка.
|
|
|
Механику перистальтики можно наблюдать в виде четких волн при бариевом исследовании пищевода. Продвижение бария по пищеводу происходит со скоростью, соответствующей манометрически измеренной скорости сокращения. Давление в пищеводе, необходимое для его эффективного опустошения, повышается по мере приближения комка к дистальному отделу пищевода. Кроме того, для эффективного прохождения твердого пищевого комка необходимо более высокое давление. И хотя жидкости в дистальных отделах пищевода проходят преимущественно под действием силы тяжести, для нормального пассажа любого пищевого комка требуется интактная перистальтическая функция.
Физиологические механизмы контроля за сокращением поперечнополосатой мускулатуры пищевода осуществляются главным образом возбуждающим действием блуждающего нерва; последовательная иннервация стимулирует нормальный процесс перистальтики (рис. 2-3). Экспериментальные исследования, в которых использовалась перерезка блуждающего нерва и реанастомоз дистального сегмента нерва с эфферентными нервами поперечнополосатой мускулатуры, показали последовательные спайк-разряды во время перистальтики поперечнополосатых отделов мышц пищевода. При прохождении через пищевод большого или твердого пищевого комка афферентные волокна, идущие от пищевода, повышают активность блуждающего нерва. Эти данные подтверждают, что поперечнополосатый мышечный слой пищевода управляется центральными нервными структурами аналогично мускулатуре верхних отделов глотки.
Перистальтика гладкой мускулатуры пищевода отличается от перистальтики поперечнополосатой за счет различий в иннервации для каждой группы мышц. В основном поперечнополосатые мышцы иннервируются блуждающим нервом без участия регуляции интрамуральными сплетениями. Регуляция сокращения поперечнополосатых мышц берет начало в центральных структурах нервной системы. Хотя центральный механизм присутствует и в регуляции гладких мышц, полная центральная денервация не прекращает перистальтику. Вагусная регуляция является важной для изменения реакции через чувствительные окончания, вызывая более сильные перистальтические волны, если пищевой комок большой или плот
|
|
|
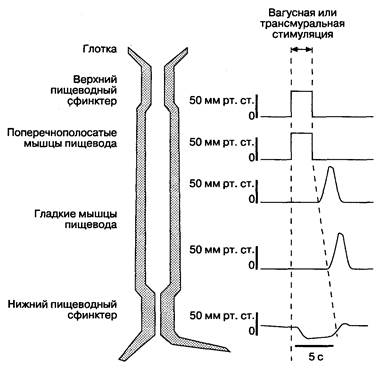
Рис. 2-3. Поперечнополосатые мышцы пищевода имеют регулируемую центральными отделами нервной системы последовательность сокращений от проксимального отдела к дистальному. Этот центральный ответ хорошо виден, когда вагусная стимуляция без учета центральной нервной системы вызывает одновременное сокращение всей поперечнополосатой мускулатуры пищевода. После прекращения вагусной стимуляции, гладкая мускулатура начинает перистальтировать. Эти данные говорят о начальном угнетении интрамуральных нейронов вагусным влиянием, за чем следует перистальтика, стимулированная либо нервами, либо мышцами, активность которых отсрочена. (По: Yamada Т., Alpers D. H., Owyang С., Powell D. W., Silverstein F. E., eds. Textbook of Gastroenterology, 2nd ed. Philadelphia:
J. B. Lippincott, 1995: 129.)
ный. Вторичная перистальтика вызывается растяжением пищевода и регулируется с помощью афферентной регуляции через блуждающий нерв, поэтому при его перерезке этот вид перистальтики нарушается.
Внутренний контроль перистальтики гладкой мускулатуры осуществляется разветвленной интрамуральной сетью. Участие интрамуральной сети в регуляции перистальтики подтверждается тем, что после перерезки блуждающего нерва перистальтика сохраняется. Эта сеть содержит как возбуждающие, так и угнетающие нейроны. Возбуждающие нейроны стимулируют сокращение продольных и циркулярных мышечных слоев через холинергические М2-рецепторы. Ингибирующие нейроны преимущественно локализуются в циркулярном мышечном слое и угнетают сокращение через нехолинергические и неадренергические нейротрансмиттеры. Основными тормозными нейротрансмиттерами считаются вазоактивный интестинальный пептид и оксид азота. Холинергическая стимуляция возбуждающих нейронов происходит через никотиновые рецепторы, в то время как нехолинергическое и неадренергическое возбуждение может осуществляться через никотиновые и мускариновые (m1) рецепторы. Оба типа нейронов иннервируют тело пищевода и нижний эзофагальный сфинктер. Более тонкие механизмы, ответственные за перистальтику пищевода, до сих пор неизвестны. Выявлено, что электрическая стимуляция пищевода вызывает вначале угнетение всей мускулатуры пищевода с последующим выраженным сокращением, напоминающим перистальтическую волну, после прекращения стимуляции. Перистальтика вызывается последовательной деполяризацией интрамуральных нейронов, распространяющейся вниз по пищеводу. Перистальтика пищевода, вызванная электрической стимуляцией, не идентична физиологической перистальтике, инициированной глотанием. При электрической стимуляции отсутствует обязательная центральная регуляция гладкой мускулатуры пищевода через блуждающий нерв, которая вызывает последовательное сокращение мускулатуры пищевода от проксимального отдела к дистальному.
Исходя из современных, знаний, можно предположить, что перистальтика пищевода регулируется преимущественно нейронально, поскольку эксперимантально выявлено, что воздействие тетродотоксином (ингибитор активности нейронов) нарушает перистальтику, кроме того, перистальтика начинается сверху вниз независимо от того, в какой части пищевода она возникла. Предполагается также возможная роль миогенной активности в пищеводе, независимой от нервной стимуляции, поскольку в мышечных препаратах, "денервированных" с помощью тетродотоксина, отмечались медленные мышечные сокращения, напоминающие спонтанную перистальтику. Однако высокочастотные стимулы, к которым восприимчива только мышечная ткань пищевода, вызывают распространение мышечных сокращений как вверх, так и вниз. Поэтому собственная мышечная активность пищевода не соответствует перистальтике, стимулированной, например, глотанием. Эти собственные свойства мышц зависят от мембранного потенциала покоя, концентрации ионов калия и проницаемости клеточной мембраны для ионов кальция. Очевидно, что спонтанная перистальтика, стимулированная глотанием, зависит как от центральной, так и от интрамуральной нервной регуляции; в свою очередь последняя модулируется собственным миогенным тонусом. Как и нервная регуляция, миогенная активность способствует более длительному мышечному сокращению гладкой мускулатуры от проксимальных к дистальным отделам пищевода.
Нижний пищеводный сфинктер (НПС) является главным барьером между кислым содержимым желудка и просветом пищевода (преимущественно щелочным). Хотя раньше предполагалось, что НПС является в основном не функциональным, а анатомическим барьером, недавние исследования выявили наличие толстого мышечного кольца, которое направлено косо вверх от малой к большой кривизне желудка. Средняя длина этого кольца составляет 31 мм. Оно соответствует зоне пищевода, в которой давление, измеренное манометрически, достигает максимальных значений. Кроме того, в нижний физиологический барьер пищевода входит часть правой ножки диафрагмы. Доказано, что сокращение правой ножки диафрагмы — главный механизм поддержки тонуса НПС при повышении давления в брюшной полости, что предотвращает обратный рефлюкс.
НПС является сегментом тонически сокращенных гладких мышц в дистальном отделе пищевода длиной 3—4 см. У здоровых людей в состоянии покоя тонус НПС составляет 10—30 мм рт. ст. Минимальное давление НПС определяется после приема пищи, максимальное — ночью. Тонус НПС преимущественно миогенный, так как он не нарушается тетродотоксином. Поскольку тонус НПС снижается под действием атропина, можно говорить и о наличии нейрональной регуляции. Кроме того, при голодании и при приеме пищи на миогенные свойства гладких мышц НПС влияют гормоны и другие вещества.
Базальный тонус НПС угнетается при глотании, когда происходит расслабление всей гладкой мускулатуры пищевода. Расслабление НПС стимулируется блуждающим нервом через преганглионарные холинергические волокна и постганглионарные нехолинергические и неадренергические нервные волокна. Механизмы, способствующие расслаблению НПС, идентичны тем, что угнетают всю гладкую мускулатуру пищевода. Нехолинергический и неадренергический нейротрансмиттер, передающий этот сигнал, еще неизвестен, но предполагается, что им могут быть вазоактивный интестинальный полипептид, пептид гистидинизолейцин и оксид азота. Учитывая, что все три вещества потенциально участвуют во многих механизмах, действие каждого из них или их сочетанное действие может быть очень важным. Роль диафрагмы в расслаблении НПС также очень важна, поскольку во время расслабления НПС волокна ножки диафрагмы не проявляют электрическую активность, в то время как купол диафрагмы сохраняет свой тонус. Рефлекс угнетения тонуса ножек диафрагмы исчезает при ваготомии.
Другой феномен был выявлен во время исследований механизмов гастроэзофагального рефлюкса. Dent и соавторы установили, что гастроэзофагальный рефлюкс имеется и при нормальном базальном тонусе НПС у здоровых людей при его транзиторном расслаблении, не связанном с глотанием. У больных с эзофагитом частота транзиторного расслабления нижнего пищеводного сфинктера, сочетающегося с кислым рефлюксом, значительно выше, чем у здоровых людей. Преходящие расслабления НПС являются главным механизмом выхода воздуха при отрыжке. Поэтому данный феномен расценивают как физиологический. Растяжение желудка сочетается с выраженным расслаблением НПС. Некоторые исследователи рассматривают транзиторное расслабление НПС как проявление неполноценного акта глотания. Доминирует точка зрения, что транзиторное расслабление НПС, не связанное с глотанием, имеет важное значение для процесса выхода газов из желудка после приема пищи.
|
|
|
