 |


Размерно-половая структура популяции
|
|
|
|
Если общее соотношение полов в популяции многих видов рыб и близко к отношению 1: 1, то в разных возрастных и размерных группах оно весьма различно. Самцы обычно преобладают в младших возрастных группах. Это связано с тем, что они созревают раньше самок, раньше вступают в состав нерестового стада и обладают меньшей продолжительностью жизни. Так, например, у камбалы Platessa platessa (L.) Северного моря (Hefford, 1903, по Wimpenny, 1953) количество самцов в 2-летнем возрасте достигает 63%, затем оно постепенно уменьшается, составляя у 5-6-летних рыб 50-51%, а у 9-летних только 13%. У речной камбалы Южной Балтики (Cieglewicz a. Mulicki, 1938) наблюдается то же: среди рыб 8-летнего возраста самцов только 20,4%, а среди 3-леток их до 83,4%. У речной камбалы восточной части Балтийского моря, по наблюдениям М. И. Шатуновского (1963), в младших возрастных группах преобладают самцы, в старших - самки. Самцы преобладают в младших возрастных группах, а самки в старших также у обыкновенного карася. Среди 3-летних карасей длиной 50-80 мм преобладают самцы, а рыбы длиной более 80 мм и в возрасте 4-5 лет представлены исключительно самками (Григораш и Спановская, 1962). У большого желтого горбыля Рseudosciaena crocea (Rich.) самцов больше среди рыб длиной менее 28 см; в группе рыб длиной 28-36 см соотношение полов равно 1: 1; у рыб крупнее 36 см больше самок (Чэнь Жу-фень и др., 1962). То же отмечено и у многих других рыб.
Однако преобладание мелких самцов в нерестовой популяции свойственно далеко не всем видам рыб. У многих рыб, например сомов (Silurus, Glyptosternum), наоборот, самки мельче самцов и их больше среди мелких рыб в нерестовой популяции.
По характеру соотношения полов среди особей разных размеров Д. Ф. Замахаев (1959) выделяет три группы рыб. У первой группы, в качестве примера которой он приводит океаническую сельдь и арктическую треску, не наблюдается заметной разницы в темпе роста, возрасте полового созревания и продолжительности жизни самцов и самок. Подобный тип соотношения полов при пространственной разобщенности нерестилищ рыб разного размера обеспечивает сохранение необходимого соотношения полов во всех размерных группах и успешное осеменение икры самок всех размеров и возрастов. Второй тип соотношения полов с размерами и возрастом характерен и для очень многих видов рыб - осетровых, большинства карповых, многих лососевидных (сиги), окуневых, камбал и др. У рыб с таким типом соотношения полов самцы становятся половозрелыми раньше самок и обычно раньше умирают. В связи с этим среди мелких молодых половозрелых рыб преобладают самцы, а среди крупных - самки (рис. 45 и 46). Преобладание самок среди крупных рыб наблюдается и у рыб в пределах одной возрастной группы. Подобная структура популяции, когда самки оказываются крупнее самцов, обеспечивает большую плодовитость стада, поскольку крупные рыбы откладывают больше икринок. В то же время это обеспечивает вследствие более мелких размеров самцов большую - при одной и той же кормовой базе - численность популяции при сохранении ее воспроизводительной способности.
|
|
|
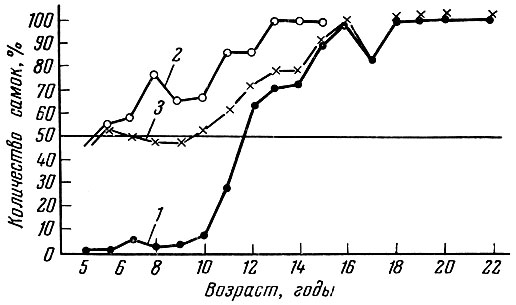
Рис. 45. Соотношение полов по возрастам у баренцевоморской камбалы (Канинская банка, 1935 и 1936 гг.) (по Замахаеву, 1959): 1 - половозрелые (18% самок); 2-неполовозрелые (62% самок); 3 - смешанная популяция (52% самок)
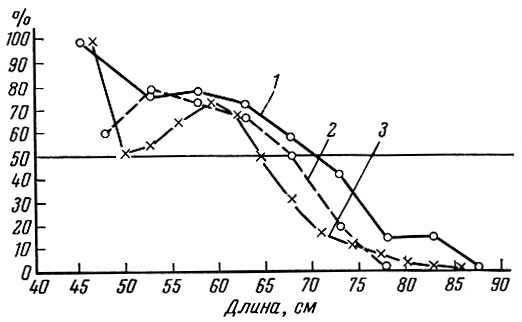
Рис. 46. Размерно-половые соотношения дальневосточных лососей (осенняя кета, р. Амур) (по Замахаеву, 1959): 1 - 2117 экз., 43% самок, 1907- 1908 гг. (по Солдатову, 1915); 2 - 1124 экз., 63,5% самок, 1937 г.; 3 - 13384 экз., 56% самок, 1940-1942 гг. (по Смирнову, 1947)
У большинства видов этой группы при нересте с более крупными самками нерестуют несколько более мелких самцов. Исключение в этом отношении представляют проходные сельди. Как показал Д. Ф. Замахаев (1959), на нерестилищах черноспинки Caspialosa kessleri (Grimm), расположенных в русле реки на течении, размерный состав самцов и самок оказывается более или менее сходным. Это, по мнению Замахаева, связано с необходимостью обладать сходными гидродинамическими свойствами (иногда, видимо, самцы, вынужденные дольше держаться на нерестилищах, должны быть сильнее самок).
|
|
|
Третья группа рыб, выделяемая Д. Ф. Замахаевым, - это виды, у которых самцы крупнее самок, и они преобладают среди рыб более крупных размеров. Иногда при этом бывает, что по возрасту соотношение полов оказывается обратным. Это наблюдается, например, у нерки Oncorhynchus легка Walb. Самцы бывают крупнее и старше самок обычно у тех видов рыб, у которых самцы охраняют икру. Это отмечено у сомов, губанов, бычков и некоторых других рыб. У проходных лососей большие размеры самцов, видимо, связаны с необходимостью преодоления ими больших скоростей течения за более длительное время (один самец нерестует с несколькими самками).
Конечно, выделенные Замахаевым (1959) три типа размерно-половых соотношений у рыб связаны между собой всей гаммой переходов. В некоторых случаях, как это отмечает в отношении сельди и сам Замахаев, у одних популяций этого вида соотношение полов первого типа, у других - ближе ко второму. То же самое, видимо, наблюдается и у трески. Однако несомненно, что намеченные Д. Ф. Замахаевым типы отражают определенные формы связей популяции со средой.
Роль карликовых самцов
У многих рыб наряду с крупными самцами, иногда по размерам превышающими самок, в популяции есть и так называемые карликовые самцы, например у лососей. Иногда самцы бывают представлены только карликовыми формами, например у глубоководных удильщиков.
У многих проходных рыб в мигрирующей части популяции процент самок бывает много выше процента самцов; это отмечено у многих лососевых, гольцов рода Salvelinus и аральского усача Barbus brachycephalus Kessl. В ходовых косяках семги очень часто преобладают самки (Берг, 1935; Азбелев и Лагунов, 1958). В некоторые годы количество самок достигает 80%. В Печоре самки составляют в иные годы 76% уловов семги, причем в верхних участках реки проходных самок может быть еще больше. На нерестилищах верховьев Печоры проходных самцов почти не бывает и в осеменении икры проходных самок участвуют карликовые самцы. На нерестилищах, расположенных ниже по течению, процент проходных самцов становится все выше (Никольский и др., 1947). Преобладают самки в уловах лосося и в бассейнах оз. Кросс в Канаде (Warner, 1962).
|
|
|
Соотношение самцов и самок лососей при проходе через счетное сооружение на р. Экз (Девоншир) равно 1: 1,5 (Allan, Swain a. Bulleid, 1960). Отмечаемый в литературе избыток самок лососей на нерестилищах - это обычно кажущееся явление и зависит оно от того, что не всегда принимаются во внимание карликовые самцы. Численность карликовых самцов может колебаться в зависимости от экологических условий размножения в разных реках (Европейцева, 1962). При искусственном выращивании молоди куринского лосося 28-30% молоди составляют карликовые самцы (Протасов и Козлова, 1957). В период массового ската молоди балтийского лосося примерно 1/4 часть молоди составляют карликовые самцы (Галкин, 1961). У мальмы Salvelinus alpinus malma на Камчатке существует обратная корреляция между численностью карликовых и проходных самцов (Савваитова, 1960). У видов лососей как Salmo, так и Oncorhynchus карликовые самцы, постоянно живущие в реках, вполне обеспечивают успешное осеменение икры крупных самок. На рыбоводных заводах их сперма может заменить сперму проходных самцов.
Иногда популяция бывает представлена двумя группами рыб обоего пола - карликовыми самцами и самками и крупными проходными самцами и самками. Например, у черноморского лосося в р. Черной, по данным Г. П. Барача (1952), среди проходных рыб 85% самок и 15% самцов, а в жилой популяции 76% самцов и 24% самок. Позднее, в 1955 г. среди проходных рыб этой реки самцов было 10% и самок 90%, а среди жилых - самцов 95% и самок 5% (Панов, 1958). Нерест у жилых и проходных рыб происходит совместно, и у молоди в речной период жизни соотношение полов близко к 1: 1. Проходные самцы лосося после нереста обычно гибнут, видимо, в большем проценте, чем карликовые, которые нерестуют не 1 раз в жизни. Многие карликовые самцы после первого нереста скатываются в море и следующий раз уже нерестуют как проходные (Митанс, 1973).
|
|
|
При сокращении численности стада проходного лосося и улучшении вследствие этого питания молоди в реке процент самок в жилой популяции возрастает.
У губанов Crenilabrus ocellatus Forsk., С. quinquemaculatus Risso, кроме крупных самцов, которые осеменяют икру и охраняют после откладки икры гнездо, есть еще мелкие самцы, которые участвуют в осеменении откладываемой в гнезде икры, но гнездо не охраняют (Soljan, 1930). Сильнее всего размеры самцов и самок различаются у глубоководных удильщиков (подотр. Ceratioidei) и у некоторых Stomiatoidei (Берг, 1937).
Гермафродитизм
У многих рыб имеется закономерный гермафродитизм. В настоящее время постоянные гермафродиты известны у представителей четырех семейств: Serranidae, Sparidae, Maenidae и Centracanthidae; все морские рыбы, главным образом тропических и субтропических широт.
Истинные гермафродиты подразделяются на: а) функциональных, или синхронных, и б) нефункциональных. У рыб первой группы в гонадах различается яичниковая и семенниковая части, и одновременно могут находиться как зрелые яйца, так и сперма. Самооплодотворения, как правило, у этих рыб не происходит, хотя у некоторых видов оно возможно. Так, Кларк (Clark, 1959) отмечает, что изолированные в аквариуме особи функционального гермафродита Serranellus subligarius откладывают оплодотворенную икру. Л. П. Салехова (1963) проводила оплодотворение икры каменного окуня Serranus scriba L. спермой той же особи и показала, что развитие икры протекает нормально, хотя лучшая выживаемость бывает при перекрестном оплодотворении. Однако в природных условиях, вероятно, самооплодотворения не происходит, и каждая особь выполняет поочередно то функцию самки, то функцию самца.
К группе синхронных гермафродитов принадлежат многие виды п/сем. Serraninae (сем. Serranidae), как Hypoplectrus unicolor (Walb.), Prionodes phoebe (Poey), P. tabacarius (Cuvier a. Valeric), P. tigrinus (Bloch) (Smith, 1959).
У рыб, принадлежащих к группе нефункциональных гермафродитов, в гонаде также различаются яичниковая и семенниковая части, однако в отличие от функциональных у нефункциональных гермафродитов обе части одновременно не функционируют. У одних видов рыб в молодом возрасте наибольшего развития достигает яичниковая часть, а самцовая остается неактивной. Такие особи функционируют как самки. По прошествии одного или нескольких икрометаний яичник у них подвергается редукции, овоциты резорбируются, а семенник развивается. Такое явление смены полов носит название протогинии. Протогиния встречается у рыб сем. Serranidae п/сем. Epinephelinae - Epinephelus guttatus (L.), E. striatus (Bloch), Mycteroperca bonaci (Poey),M. tigris (Cuvier a. Valenc) (Smith, 1959),
|
|
|
сем. Maenidae - Pagellus erythrinus (Zei a. Zupanovic, 1961), сем. Sparidae - Diplodus annularis L., D. sargus (L.) и др. (D'Ancona, 1950), Taius tumifrons (Aoyama, 1955), у представителя сем. Centracanthidae - Spicara maena (Reinboth, 1962).
У других видов наблюдается иная закономерность. В младших возрастах они функционируют как самцы, а в старших как самки. Такая смена полов носит название протандрии. Протандрия наблюдалась у рыб сем. Sparidae - Diplodus sargus, Pagellus mormyrus (D'Ancona, 1950), Diplodus annularis (Салехова, 1961; Reinboth, 1962).
По данным Л. П. Салеховой (1961), в популяции Diplodus annularis есть как раздельнополые особи, так и гермафродиты, причем процент гермафродитных особей уменьшается с возрастом. Так, среди 4-летних рыб самок 60/6, самцов 20% и гермафродитов 20%, а среди 6-летних гермафродитов всего 3%.
Третий тип потенциального гермафродитизма известен у рыб сем. Labridae (Bacci a. Razzanti, 1957; Reinboth, 1961,1962; Okada, 1962; Sordi, 1961, 1962) и у одного представителя отряда Symbranchiformes (Lin, 1944; Liem, 1963). Рыбы этой группы в младших возрастах имеют яичник и функционируют как самки, затем у них происходит реверсия пола, и на старших возрастах рыбы представлены только самцами. Здесь также наблюдается протогиния, но в отличие от протогинии нефункциональных гермафродитов у рыб с потенциальным гермафродитизмом в гонаде самки нет выраженной семенниковой части. Среди овоцитов есть лишь недифференцированные половые клетки - гонии, дальнейшее развитие которых и образует семенник взамен яичника.
У Coris julis Günth. наблюдаются краткая фаза смены пола и длительные периоды пребывания самками или самцами. Превращение пола у этого вида связано с изменением окраски, отчего молодых рыб Coris julis, находившихся в периоде женского развития, принимали раньше за другой вид - Coris giofredi Risso. Monopterus albus (Zuiev) обитает на рисовых полях Юго-Восточной Азии и Малайского архипелага. У этого вида молодые особи функционируют как самки, старые-как самцы. Общее соотношение полов: 3 самки на 1 самца.
Биологический смысл закономерного гермафродитизма не всегда еще ясен. Протогиния и соотношение полов, сдвинутое в сторону преобладания самок на младших возрастах, видимо, есть приспособление для увеличения скорости воспроизводства в крайне неблагоприятных условиях. Так, Monopterus albus обладает коротким периодом размножения, и он сумел приспособиться к крайне нестабильным условиям жизни на рисовых полях. После сухого периода популяция быстро восстанавливается благодаря тому, что все кормовые ресурсы используются в первую очередь для обеспечения жизни особей, функционирующих как самки; это приводит к повышению численности популяции. Пережившие неблагоприятный период особи становятся самцами. Эти рыбы крупнее, они продуцируют больше спермы и способны нерестовать с несколькими самками.
Таким образом, протогиния - приспособление, обеспечивающее увеличение численности популяции путем использования кормовых ресурсов водоема особями, функционирующими как самки. Протогиния, очевидно, присуща видам, живущим в таких водоемах или частях водоемов, где наблюдаются нестабильные условия и где ограничена кормовая база.
Протандрия встречается несколько реже. Она, очевидно, связана с тем, что более мелкие особи, функционирующие как самцы, в состоянии обеспечить успешное осеменение икры, а более крупные особи, становящиеся самками, повышают плодовитость популяции.
В случае синхронных гермафродитов обеспечивается возможность восстановления популяции даже при сохранении единичных особей.
Строгое распределение гермафродитных видов по указанным группам гермафродитов не всегда возможно. Детальное изучение развития гонад в течение нескольких лет на одном поколении морского карася Diplodus annularis, проведенное Л. П. Салеховой (1961), показало, что у этого вида встречаются и раздельнополые особи, и функциональные гермафродиты и наблюдается протандрия. Таким образом, диапазон половых изменений у этого вида гораздо шире, чем указывали ранее другие исследователи. У старых рыб в связи с угасанием половой функции также может наблюдаться реверсия пола, что было отмечено у самок гольяна, карпов, речной камбалы (Анисимова, 1956, 1956а; Bullough, 1940; Микельсаар, 1958; наблюдения А. П. Макеевой).
|
|
|
