 |


Приспособительное значение изменений соотношения полов и механизмы регуляции половой структуры популяции
|
|
|
|
Поскольку половая структура популяции имеет очень важное значение в воспроизводстве стада, то естественно, что у рыб, как и у других животных, вырабатываются механизмы регуляции полового состава стада в связи с меняющимися условиями жизни, в первую очередь с изменениями обеспеченности пищей. Так как обеспеченность пищей особей тесно связана с численностью кормящейся популяции, то естественно, что изменения численности должны отражаться определенным образом на соотношении полов в популяции, меняя ее воспроизводительную способность.
В настоящее время накоплен уже большой материал, говорящий о связи изменений соотношения полов и численности популяции у рыб и круглоротых. Так, у ручьевой миноги Lampetra planeri (Bloch) в реках Англии, по данным Хардисти (Hardisty, 1954), процент самок в популяции в р. Иео (Jeo, Англия) определяется общей численностью популяции и он тем ниже, чем выше численность популяции. Последующие наблюдения Хардисти (1960) еще с большей четкостью показали, что чем больше численность популяции ручьевой миноги, тем больше в ней самцов. Сходная закономерность отмечена и у пресноводной формы миноги Petromyzon marinus L. бассейна Великих Озер Северной Америки (Applegate, 1950), где в годы с более высокой численностью популяции самцов больше самок (табл. 73).

Таблица 73. Изменение соотношения полов у миноги англии в связи с изменением численности (по Hardisty, 1954)
Такая же закономерность отмечена у ряда видов рыб, в частности у амурской горбуши. М. Л. Крыхтин и А. Г. Смирнов (1962) указывают, что "в малочисленных нерестовых стадах горбуши самки в течение многих лет преобладали лад самцами, а в годы сравнительно высокой численности (1948 и 1958), наоборот, самцов было заметно больше, чем самок".
|
|
|
В отношении горбуши Южного Сахалина Г. М. Персов (1964) установил, что в годы, когда молодь горбуши мельче, у нее преобладают самцы (нечетные годы) и, наоборот, в годы, когда молодь крупнее, возрастает процент самок. Об увеличении количества самцов в многочисленных популяциях озерновской красной любезно сообщила нам Т. В. Егорова. Увеличение количества самцов ротана Percottus glehni Dyb. до 60% при ухудшении условий жизни в результате значительного увеличения численности популяции наблюдали В. Д. Спановская, К. А. Савваитова и Т. Л. Потапова (1964).
Обеспеченность популяции пищей и качество этой пищи являются главными причинами изменения соотношения полов как у рыб, так и у других животных. При лучших условиях откорма соотношение сдвигается в пользу самок, при худших - самцов. По данным Н. С. Олейникова, Л. И. Широковой и Н. Н. Павловой (1957), в оз. Абрау численность рыб по ряду причин заметно снизилась, в связи с чем кормовая база стала недоиспользоваться рыбой. Благодаря улучшению кормовой базы количество самок в популяциях красноперки возросло по сравнению с самцами на 12,9%, а самок у сазана стало в 1,5 раза больше, чем самцов.
Регулярно наблюдается некоторое преобладание самцов у подуста в реках Трансильвании (Szabo, 1958), у обыкновенного карася в некоторых озерах бассейна Печоры (Никольский и др., 1947). В этих случаях увеличение количества самцов связано, по-видимому, с бедной кормовой базой.
Крумхольц (Krumholz, 1963) в течение 16 месяцев изучал популяции гамбузии из мангровых зарослей и солоноватого пруда, где не было хищников. Половая зрелость у рыб из мангровых зарослей наступает при меньших размерах и они плодовитее, чем гамбузии в пруду. Среди рыб мангровых зарослей преобладают самки, в пруду - самцы, есть также старые самки. То, что недостаток пищи, часто являющийся следствием высокой численности популяции, обычно приводит к увеличению количества самцов, подтверждается и экспериментами. Так, закономерное изменение соотношения полов Эберхардт (Eberhardt, 1943) установил на примере бойцовой рыбки Betta. Он показал, что ухудшение обеспеченности ее пищей вызывает изменение соотношения полов по сравнению с типичным 1: 1 в сторону увеличения процента самцов. Д'Анкона (D'Аnкоnа, 1950) экспериментально показал, что выращивание личинок угря при плотных посадках вызывает преобладание среди них самцов. Д'Анкона приводит аналогичные данные других исследователей - Теша (в Голландии), Гондолеи и Хорнхольда (во Франции), выращивавших личинок угря в разных условиях.
|
|
|
Из приведенных примеров видно, что увеличение плотности популяции, снижая обеспеченность рыб пищей, увеличивает количество самцов в популяции. Но у некоторых видов рыб снижение обеспеченности пищей, наоборот, вызывает появление большого количества мелких самок. Эти самки рано созревают и способны поддерживать высокую численность популяции даже в неблагоприятных условиях. С таким явлением мы встречаемся, в частности, при изучении обыкновенного окуня. Соотношение полов у этого вида варьирует в самых широких пределах. В одних вэдоемах преобладают самки: в оз. Зайсан (Дианов, 1956), в Малом Море Байкала и оз. Заглинур (Рекстин, 1959; Евтюхова-Рекстин, 1952), в Байкале (Асхаев, 1958), в бассейне р. Печоры (Никольский и др., 1947), в Москве-реке (Григораш и Спановская, 1962), в Онежском озере (Гуляева, 1951), в оз. Вакшти (Соловьева, 1960); в других водоемах преобладают самцы: в оз. Синтур в Сибири (Пробатов, 1936), в оз. Вильгортском (Соловьева, 1960), в водоемах Ленинградской области (данные, любезно сообщенные нам Л. А. Жаковым); в третьих водоемах соотношение полов почти равное: в бассейне р. Кети (Мирошниченко и Суханова, 1952), в оз. Убинском в Западной Сибири (Никольский, 1953). Количество самок может значительно превосходить количество самцов и достигать 9 самок на 1 самца. Альм (Aim, 1959) предполагает, что в некоторых случаях изменение в соотношении полов у окуня обусловливается селекцией орудий лова. Однако основная причина изменений в соотношении полов у окуня - это результат разной обеспеченности пищей (Dyk, 1938). В условиях плохого питания происходит также выделение двух форм окуня - быстро и медленно растущих, и многочисленные мелкие тугорослые самки служат пищей более крупным самкам. Кроме того, у обыкновенного окуня в некоторых популяциях наряду с крупными самцами появляются и карликовые самцы, становящиеся половозрелыми в годовалом возрасте (Шарашидзе, 1968; Спановская, Григораш и Лягина, 1973). Эти самцы участвуют в нересте с крупными самками. Карликовые самцы появляются в популяциях окуня спорадически в условиях высокой обеспеченности пищей. Так как из-за малых размеров они плохо улавливаются орудиями лова, то, возможно, что малый процент самцов в некоторых популяциях окуня является результатом недоучета карликовых самцов.
|
|
|
Интересные данные получены о камбале-глоссе Хаджибейского лимана Черного моря (Замбриборщ, 1956). У этого вида в условиях лимана также значительно преобладают самки. Они рано созревают и очень плодовиты. Автор справедливо рассматривает этот факт как ответ популяции на неблагоприятные условия жизни в лимане. Линдсей (Lindsey, 1962), инкубируя икру трехиглой колюшки при одинаковой температуре и солености, но при разной плотности посадки, отметил, что при большей плотности посадки возрастает количество самок.
Оказалось, что и в потомстве рыб, живших в переуплотненных популяциях, самки могут превосходить по количеству самцов; это установлено у Lebistes (Breder a. Coates, 1932).
В ряде случаев изменение соотношения полов у разных размерных и возрастных групп в популяции также связано с изменением численности поколений и изменениями обеспеченности пищей.
Изменение мощности поколения, вступающего в стадо, может сильно изменить соотношение полов в отдельных размерных группах. Сначала резко возрастает процент самцов среди младших и самых мелких производителей, а затем увеличивается относительное количество позднее созревающих самок. Так, например, в мощных поколениях севрюги, появившихся в результате запуска времен гражданской войны, впервые годы резко увеличилось количество самцов. Отметим, что у севрюги, несомненно, состав уловов отражает состав нерестового стада (табл. 74).
|
|
|
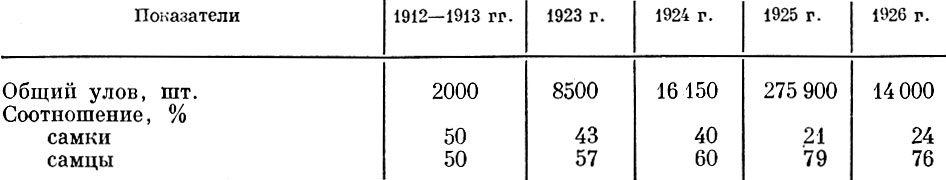
Таблица 74. Общий улов и соотношение полов у кубанской севрюги в районе Ачуева (по Чугунову, 1927)
Н. Л. Чугуиов (1927) считает, что "преобладание самцов севрюги в промысловых уловах 1925 и 1926 гг. - результат непосредственного влияния периода запуска рыболовства, так эта волна является первым половозрелым поколением самцов приплода 1919-1920 гг.". Вполне возможно, что подобного рода изменения в соотношении полов могут произойти и в результате изменения естественной смертности или изменения обеспеченности пищей вследствие изменения кормовой базы или условий нагула.
Сходную картину у дальневосточных лососей отмечает Матисен (Маthisen, 1955, 1962): самки нерки Oncorhynchus nerka Walb. становятся половозрелыми в более старшем возрасте, чем самцы. То же наблюдается у чавычи О. tschawytscha (Walb.).
Когда в половозрелое стадо вступает мощное пополнение, оно изменяет соотношение полов в пользу самцов, поскольку самцы созревают раньше самок. К. И. Мишарин (1958) указывает, что в годы, когда вступает мощное поколение байкальского омуля, количество самцов сильно увеличивается и может составлять более двух самцов на одну самку. Обычное соотношение полов у омуля - 34% самок и 66% самцов - автор считает наиболее благоприятным для естественного размножения. Такое соотношение полов наблюдается в стаде, не нарушенном промыслом. Сходное явление отмечено и у других рыб, в частности у плотвы Можайского водохранилища (Спановская, Григораш и Лягина, 1963). В первый год залития Можайского водохранилища улучшились условия откорма рыб и ускорилось развитие их половых желез. На второй год залития в нерестовой популяции плотвы количество самцов, созревающих раньше самок, увеличилось до 59 вместо 23,7%. Более сложная картина наблюдается, если с изменением плотности популяции, в частности мощности поколений, меняется темп полового созревания. Если изменение возраста полового созревания у обоих полов носит сходный характер, соотношение полов в разных размерных и возрастных категориях не изменяется. Если же есть разница в сдвиге темпа полового созревания у самцов и самок, то это, отражается на изменении размерно-половой структуры популяции.
Так, например, у обыкновенного карася Carassius carassius (L.) в исключительно благоприятных условиях роста разница во времени созревания самцов и самок сглаживается (в обычных условиях самцы созревают несколько раньше самок) и оба пола созревают в одном возрасте 1+(Schäperclaus, 1953). При этом относительная численность самцов уменьшается среди младших возрастных групп, но возрастает среди старших.
|
|
|
Соотношение полов в нерестующей части стада может существенно изменяться из-за пропуска самками нереста в неблагоприятные годы.
Так, например, в годы с неблагоприятными условиями нагула или в поколениях высокой численности у беломорской сельди Clupea harengus maris-albi Berg часть самок нерестует не каждый год и, естественно, при этом возрастает процент самцов на нерестилищах (Чепракова, 1966). У сигов Coregonus lavaretus (L.) (Решетников, 1966), у леща в северной части его ареала (Кошелев, 1966) и некоторых других рыб самки участвуют в нересте не ежегодно, что также, естественно, отражается на соотношении полов на нерестилищах. Сходную картину отметили у чира р. Рыбной (бассейн р. Пясины) В. А. Красикова и С. М. Сесягин (1962) и у чукучанаоз. Невольничьего Северной Америки Рой (Roy, 1962) и др.
У млекопитающих установлена закономерная связь возраста родителей и соотношения полов в потомстве (Курбатов, 1954). О рыбах таких данных очень мало. Нам удалось обнаружить только данные о бойцовой рыбке (табл. 75). У этого вида при скрещивании рыб старшего возраста процент самцов в потомстве возрастает, что также, видимо, является одним из регуляторных механизмов популяции.

Таблица 75. Изменение соотношения полов в потомстве ветта Splendens в связи с возрастом родителей (по Eberhardt, 1943)
По данным Тумма (Thumm, 1908,) при скрещивании крупных 3-годо-валых самок Cichlasoma nigrofasciatum Günth. с мелкими годовалыми самцами в потомстве преобладали самцы (750 самцов на 50 самок). При обратном скрещивании старых самцов с молодыми самками - соотношение полов было обратным (на 100 самцов 300 самок).
Возможно, что в некоторых случаях может иметь место повышенная смертность одного пола до достижения половой зрелости. Это, например, указывается Шумахером (Schoemacher, 1944) в отношении Lebistes, в популяции которой соотношение полов у молоди составляло 1: 1, а через год стало 1: 4 в пользу самок вследствие повышенной гибели самцов. Однако в большинстве рассмотренных случаев изменение в соотношении полов не зависит от повышенной смертности особей одного пола.
Своеобразный и плохо изученный механизм регуляции пола наблюдается у Anthias squammipinnis. Молодые особи этого вида способны функционировать как самки, а в более старшем возрасте как самцы. Если несколько самок (10-20 шт.) держали в аквариуме вместе с одним-двумя самцами, все самки оставались самками, их поведение и строение не менялись. Если же группу самок изолировали от самцов, то через две недели у одной из них начинались изменения, появлялась характерная для самцов окраска, менялось поведение, дегенерировали яичники. И в природе, если у этого вида число самцов уменьшается ниже определенной нормы, часть самок превращается в самцов (Фишельзон, 1971).
Нарушение соотношения полов в нерестовом стаде, видимо, обычно не вызывает изменения соотношения полов в потомстве. Так, Матисен (Mathisen, 1955) указывает, что в нерестовом стаде нерки Бристольского залива в результате отбора жаберными сетями с 5 1/2 дюймов оказывается часто вдвое больше самок, чем самцов. Однако у скатывающихся из реки смолтов - потомства этих производителей с нарушенным соотношением полов - соотношение полов близко к 1: 1.
Таким образом, на изменение обеспеченности пищей популяция рыб реагирует изменением соотношения полов и размерно-половой структуры. Эти изменения могут происходить в результате как трансформации пола у рыб, так и разновременности полового созревания самцов и самок или пропуска нерестового сезона обычно самками при ухудшении обеспеченности пищей, либо в результате более интенсивной гибели одного из полов.
Несомненно, что у рыб механизмы, регулирующие половую структуру популяции, многообразны. Однако ни разницей в возрасте наступления половой зрелости, ни повышенной смертностью особей одного пола на ранних стадиях развития нельзя объяснить многие случаи изменения соотношения полов. Несомненно, что у рыб соотношение полов регулируется через изменение хода обмена, вызываемого изменением обеспеченности пищей, которое часто происходит в результате изменения плотности популяции. Это указывает на то, что соотношение полов в популяции регулируется через изменение хода обменавеществ.
Возможно, иногда регулирование пола идет через нервно-гуморальную систему. "Сигнал" поступает в организм через тот или иной рецептор.
Схематически основной механизм регуляции пола можно изобразить в виде следующей схемы.

|
|
|
