 |


Морфологическая разнокачественность особей в популяции
|
|
|
|
Морфологическая разнокачественность особей в популяции является одним из важнейших видовых приспособлений, связанных с динамикой численности и биомассы стада. Морфологические различия особей, находящихся на разных этапах онтогенеза, обеспечивают популяции освоение более разнообразных кормов и выработку специфических приспособлений к защите от хищников. Но разнокачественность особей в популяции не сводится только к возрастным различиям. И в пределах одновозрастной популяции особи оказываются морфологически различными. У многих рыб наблюдаются иногда значительные различия между самцами и самками. Это, как мы видели выше, связано, в частности, и с различиями в характере питания полов. Но и в пределах одного пола между особями в поколении есть закономерные различия, которые, как мы показали ранее (Никольский, 1955а; Никольский и Пикулева, 1958), имеют приспособительное значение и позволяют особям популяции в целом осваивать более широкий ассортимент кормов. В условиях низкой обеспеченности пищей, как показано выше, сиги с более короткими и редкими тычинками питаются одними кормами, а особи с более частыми и длинными тычинками - другими. Сходная картина в условиях низкой обеспеченности пищей отмечена нами у короткоусых и длинноусых особей пескарей.
Таким образом, морфологическая разнокачественность особей обеспечивает популяции расширение спектра ее питания. В настоящее время существуют две точки зрения на значение разнокачественности в жизни вида. По одной точке зрения, разнокачественность особей в популяции приводит к тому, что особи, оказавшиеся более приспособленными, находятся в более благоприятных условиях жизни и постепенно вытесняют менее приспособленных, которые вымирают. Этой точки зрения придерживался Дарвин.
|
|
|
По другой точке зрения, разнокачественность особей в популяции- это видовое приспособление, обеспечивающее выживание особей и сохранение популяции как относительной морфобиологической стабильности. Если правильна первая точка зрения, то различающиеся по строению особи должны отличаться друг от друга по жирности и степени развития гонад. Если правильна вторая точка зрения, то такой закономерной разницы в жирности и развитии гонад у рыб с разной степенью развития того или иного признака, связанного с питанием, обычно быть не должно.
Чтобы выяснить этот вопрос, мы попытались сопоставить жирность (масса печени в процентах от массы тела) и коэффициент зрелости гонад у самцов и самок беломорской трески с разной относительной длиной усиков. Были взяты как смешанные по возрастному составу пробы, так и отдельно особи в возрасте 4+. Во всех случаях у трески не удалось обнаружить достоверной корреляции между степенью развития усика и жирностью и степенью развития гонад (рис. 47). Хотя некоторая разница в питании рыб с разной длиной усика и наблюдается, но она не создает в обычных условиях жизни популяции преимущества для рыб, обладающих той или иной величиной этого признака. Конечно, в тех случаях, когда вид корма, к питанию которым адаптированы более длинноусые или более короткоусые особи, сильно уменьшается или полностью исчезает, эти особи могут оказаться в менее благоприятных условиях обеспеченности пищей и стать более худыми и даже частично или полностью вымереть. В оптимальных же условиях питания морфологическая разнокачественность не приводит к ухудшению условий жизни части особей, но обеспечивает более широкий спектр потребляемых популяцией кормов.

Рис. 47. Соотношение длины усика, массы печени и массы гонад у трески
|
|
|
Разнокачественность особей в поколении возникает еще на стадии икринки. Выше был рассмотрен механизм возникновения разнока-чественности в размерах икринок и количестве в них жира и показано приспособительное значение этого явления. У ряда рыб, нерестующих на гальке (осетровые, литофильные карповые), выметываемая икра даже у одной особи обладает разной клейкостью, что обеспечивает ее различный разнос течением и мешает вредной концентрации большого числа икринок в одном месте. У весенненерестующих стад осетра икра оказывается несколько менее клейкой, чем у летненерестующих. У иссык-кульского чебачка Leuciscus bergi Kasehk. в разные периоды хода икра также обладает разной клейкостью, что способствует более равномерному распределению икры на нерестовых площадях.
Поскольку количество сегментов тела, число позвонков, число чешуи в боковой линии и лучей в непарных плавниках у рыб меняется в известных пределах в зависимости от температуры развития, то естественно, что у особей, развивающихся при более низкой температуре из икринок, отложенных в начале нереста, у весенненерестующих рыб оказывается несколько большее число метамеров тела, позвонков и лучей в плавниках, чем у особей, развивающихся при более высоких температурах. Подобное явление отмечено у очень многих видов рыб: сельдей (Hempel a. Blaxter, 1961), лососей (Täning, 1952), карпозубых (Gabriel, 1944) и многих других. У сельди эта связь выражена очень четко (рис. 48). Меняются в зависимости от температуры количество метамеров тела и связанные с этим величины меристических признаков и у рыб, принадлежащих к разным поколениям. Так, М. И. Шатуновский (1964) на примере речной камбалы Кандалакшского залива Белого моря показал, что существует корреляция между поверхностными температурами воды в мае - июле и средними значениями меристических признаков (рис. 49).
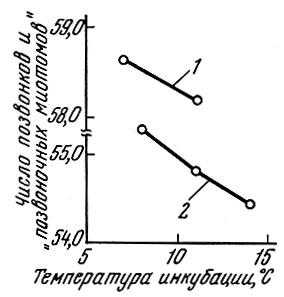
Рис. 48. Влияние температуры инкубации на число позвонков и на числа позвоночных миотомов (по Hempel a. Blaxter, 1961): 1 - в районе Клайд; 2 - в районе побережья ГДР и ФРГ
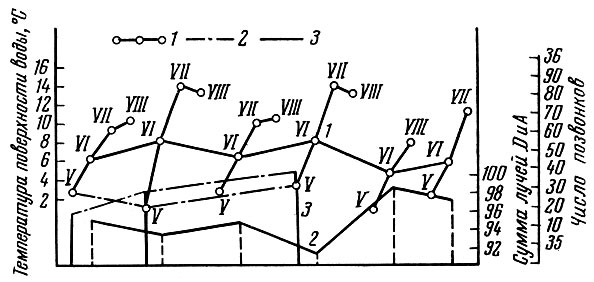
Рис. 49. Зависимость числа позвонков и лучей в спинном и анальном плавниках речной камбалы от температуры поверхностного слоя воды в мае - июле (по Шатуновскому, 1964): 1 - средняя месячная температура поверхности воды в мае - июле; 2 - суммарное число лучей в спинном и анальном плавниках камбалы; 3 - число позвонков
|
|
|
Таким образом, формируется определенная морфологическая разнокачественность в меристических признаках особей в популяции.
Однако разнокачественность особей в популяции возникает не только на стадиях метамеризации тела. По ряду признаков эта разнокачественность формируется позднее. Так, у балтийской салаки количество пилорических придатков устанавливается на четвертом - пятом этапе формирования кишечника и связано в некоторой степени с характером потребляемой пищи (Чепурнов, 1961). Еще позднее у некоторых видов рыб, например у калифорнийской сардины, заканчивается формирование числа жаберных тычинок. Таким образом, даже по меристическим признакам разнокачественность популяции формируется и на довольно поздних этапах онтогенеза.
Сводка о связи изменений меристических признаков и условий жизни сельди сделана Блекстером и Холлидай (Blaxter a. Holliday, 1963). Менее изучены изменения разнокачественности пластических признаков в популяции.
У головешки-ротана Percottus glehni Dyb. в популяции, акклиматизировавшейся в пруду под Москвой, в результате переуплотнения популяции и перехода на каннибализм изменились пропорции головы, высота тела, длина брюшных и второго спинного плавников. При этом, как указывалось выше, сильно изменился и коэффициент вариации признаков. В разреженной популяции ротана в первый период акклиматизации средний коэффициент вариации был 6,9, а во второй период при замедлении роста и высокой численности популяции он увеличился до 9,7. Интересно, что у исходной амурской популяции из района Болони средний коэффициент вариации пластических признаков равен 7,3. Изменения в высоте тела и плавниках связаны с ростом рыбы (Спановская, Савваитова и Потапова, 1964). Изменения в пропорциях головы, связанные с переходом на хищное питание, могут быть и в разреженных популяциях, как это отмечено у севанской форели, пересаженной в Иссык-Куль (Лужин, 1956), но в этом случае коэффициент вариации не увеличивается.
|
|
|
Существенно изменилась величина ряда признаков у плотвы при образовании Можайского водохранилища. Впервые годы образования этого водохранилища обеспеченность пищей стада плотвы, попавшего сюда из Москвы-реки, резко увеличилась (Лягина, 1967). В результате у плотвы сильно уменьшились относительная величина головы, высота спинного и анального плавников, длина грудных плавников (рис. 50). Относительно увеличились наибольшая высота тела, пектовентральное расстояние, длина хвостового стебля и ряд других признаков, т. е. в результате изменения обеспеченности пищей очень сильно изменяется экстерьер рыб. У плотвы это произошло вследствие резкого увеличения обеспеченности пищей в результате увеличения кормовой биомассы и нагульных площадей. Строение головешки-ротана изменилось в результате ухудшения обеспеченности пищей, вызванного увеличением численности популяции.

Рис. 50. Изменение морфологических признаков плотвы разного возраста в реке и водохранилище (по Лягиной, 1967): а - длина головы; б - постдорсальное расстояние; в - длина хвостового стебля; г - наибольшая высота тела; д - высота спинного плавника; е - расстояние между основаниями Р и V; ж - высота анального плавника; 1 - 5-летки (река), 6- и 7-летки (водохранилище); 2- 6-летки (река), 7- и 8-летки (водохранилище); 3- 7-летки (река), 8-летки (водохранилище); 4- 5-, 6- и 7-летки (1959 г., возрастные изменения в реке)
Лещ Аральского моря под влиянием изменений абиотических и биотических условий его жизни, произошедших в этом водоеме, претерпел значительные морфологические изменения. У него увеличилась относительная длина головы, уменьшилась высота тела. Сильно замедлился темп роста и снизилась упитанность и жирность. По своим признакам аральский лещ стал сходным с тугорослыми озерными формами (Борисов и Дорошев, 1970).
Во всех рассмотренных случаях изменения признаков произошли в соответствии с изменившимися условиями и являются приспособлением к этим условиям.
Заключение
Таким образом, структура популяции, как и другие видовые признаки, имеет приспособительное значение, обеспечивая существование популяции в определенных условиях. Структура популяции в известных пределах, специфичных для вида, приспособительно меняется в соответствии с изменениями условий жизни. Разнокачественность особей в популяции - закономерное явление, обеспечивающее существование вида. Однако существует и другая точка зрения, по которой структура популяции, в частности ее возрастной состав, не имеет приспособительного значения, а в значительной степени является результатом случайных внешних воздействий; разнокачественность особей в популяции - это результат или случайных внешних воздействий или реализации независимых от среды внутренних свойств родительских особей, она подчиняется только статистическим закономерностям и не имеет приспособительного значения для жизни вида, служит только субстратом для непрерывно действующего естественного отбора. Мне эта точка зрения представляется неверной. Основные закономерности, которым подчиняются изменения структуры популяции, следующие.
|
|
|
1. Средний и предельный возраст хищных рыб обычно выше, чем бенто- и планктонофагов. Самая короткая продолжительность жизни и наименьшие размеры свойственны обычно планктоноядным рыбам.
2. Продолжительность жизни видов рыб низких широт в среднем меньше, чем у видов рыб высоких широт. В пределах одного вида более южные популяции (в северном полушарии) обычно характеризуются более коротким жизненным циклом, чем более северные. У жилых популяций жизненный цикл обычно короче, чем у проходных.
3. Изменения возрастного состава, происходящие в пределах одной популяции, - это результат взаимодействия трех процессов: пополнения, роста и убыли.
4. Стабильность и изменчивость структуры популяции в значительной степени зависят от колебания урожайности отдельных поколений. У видов со значительными флюктуациями изменчивость возрастного состава популяции много сильнее, чем у видов, у которых урожайность поколений меньше колеблется по годам.
5. Возрастная структура популяции - это видовое приспособление, обеспечивающее популяции существование в конкретных условиях. Многовозрастная структура популяции есть приспособление к относительно стабильной кормовой базе, слабому воздействию хищников на половозрелую часть стада и лабильным условиям воспроизводства. Рыбы с коротким жизненным циклом и простой структурой популяции приспособлены к относительно стабильным условиям воспроизводства, лабильной кормовой базе и изменчивому и интенсивному воздействию хищников.
6. Приспособительная перестройка структуры популяции осуществляется автоматически через регуляцию роста, возраста полового созревания, продолжительности жизни и соотношения пополнения и остатка.
7. Соотношение полов в нерестовом стаде, размерно-половая структура популяции - это видовое приспособление, обеспечивающее необходимую эффективность размножения в связи с меняющимися условиями жизни.
8. В соответствии с изменениями условий жизни, в первую очередь обеспеченности пищей, меняется и половая структура популяции, в связи с чем меняется воспроизводительная способность стада.
9. Изменение половой структуры популяции осуществляется через перестройку обмена веществ и гормональной активности.
10. Морфологическая разнокачественность особей в популяции обеспечивает популяции более широкое освоение условий жизни, в первую очередь повышение обеспеченности пищей.
11. На изменения обеспеченности пищей и других условий жизни популяция приспособительно отвечает изменением амплитуды разнокачественности особей.
12. Изменения обеспеченности пищей связаны с изменениями и средних величин пластических и меристических признаков в зависимости от условий жизни.
|
|
|
