 |


Причины флюктуации численности рыб
|
|
|
|
У всех рыб, как и у большинства других позвоночных, основная гибель обычно происходит на ранних этапах развития. Разным выживанием в значительной степени определяются и колебания уловов в тех случаях, когда промысел достиг достаточной интенсивности, а иногда даже при весьма слабой интенсивности промысла урожаи и неурожаи поколений оказываются очень заметными. Так, катастрофическое падение численности стад дальневосточных лососей еще задолго до развития их интенсивного промысла приводило к голоду местного населения на Камчатке (Слюнин, 1900). Падение численности сельди по побережью Скандинавии отмечалось еще в средние века (Rollefsen, 1949, 1960). В настоящее же время при высокой интенсивности промысла урожайность поколений очень сильно влияет на продуктивность промысла.
Промысел сельди в Северо-Восточной Атлантике за последние годы в значительной степени базировался на поколении 1950 г. рождения. Почти около десяти лет промысел сахалино-хоккайдской сельди опирался на поколение 1939 г. рождения. Многочисленное поколение сельди дает промыслу в десятки раз больше рыбы, чем малочисленное. Чем меньше возрастных групп образуют нерестовую популяцию вида, тем сильнее на ее общей численности сказываются колебания урожайности отдельных лет. Лучше всего это можно проследить на примере сардин, уловы которых как в Атлантическом, так и в Тихом океане колебались только за последние полвека в сотни раз. В то же время уловы некоторых других видов рыб остаются удивительно стабильными. Хорошим примером этого могут служить уловы палтуса Hippoglossus hippoglossus stenolepis Schmidt no Тихоокеанскому побережью Канады и США. За период с 1915 по 1945 г. максимальный улов этой рыбы превысил минимальный всего примерно в 21/2раза (Thompson, 1950).
|
|
|
Флюктуации - колебания численности поколений различных годов рождения - выражены у разных видов рыб далеко не в одинаковой степени. У одних видов, например у сельди, трески Северной Атлантики (Hjort, 1914), урожайное поколение может превышать слабое во много десятков раз. Сходная картина наблюдается у сельди и трески северной части Тихого океана. То же самое, правда, с несколько меньшей амплитудой имеет место у пикши. В пределах того же бореального фаунистического комплекса как в Северной Атлантике, так и в северной части Тихого океана мы встречаем виды рыб и с очень незначительными флюктуациями. Это уже упоминавшийся палтус Hippoglossus hippoglossus (L.), морская камбала Platessa platessa (L.) и ряд других.
Рыбы со значительными флюктуациями встречаются и в пределах других фаунистических комплексов, например сардина Sardina pilchardus (L.) и пеламида Pelamys sarda (L.). В пределах средиземноморского комплекса флюктуации обычно сильно развиты у планктоноядных рыб и у питающихся этими рыбами хищников.
Таким образом, величина флюктуации, естественно в определенных пределах, специфична для вида и является его видовым свойством (Никольский, 1958а, 1961а).
Всякое видовое свойство есть приспособление, обеспечивающее сохранение и процветание вида. Флюктуации - это одна из сторон типа динамики стада. Как тип динамики популяции в целом, так и флюктуации как одна из его сторон являются видовым свойством, следовательно, они имеют для вида приспособительное значение.
Что общего между рыбами с резко выраженными флюктуациями? Флюктуации могут быть у рыб, принадлежащих как к различным систематическим группам, так и различных по своему географическому происхождению. Значительные флюктуации наблюдаются лишь у тех видов рыб, у которых количество икры, продуцируемое одной самкой в год, не очень мало.
Однако сама по себе величина плодовитости не является причиной флюктуации. Как я показал ранее (Никольский, 1961а), среди рыб и со значительными и со слабо выраженными флюктуациями мы находим рыб как с высокой, так и с низкой плодовитостью. В то же время у рыб со значительными флюктуациями и рыб, у которых флюктуации не выражены, разница в плодовитости может быть очень велика (в десятки и сотни раз). Плодовитость атлантической сельди во многие десятки, а то и в сотни раз ниже плодовитости трески, амплитуда же флюктуации у этих видов очень близка. Таким образом, прямой связи между плодовитостью вида и флюктуациями величины его поколений не наблюдается. По-видимому, флюктуации у рыб так же, как это отмечено многими авторами в отношении наземных позвоночных и насекомых, как-то связаны с обеспеченностью пищей.
|
|
|
Такие виды, как треска, сельдь, пикша, сардины, снеток, приспособлены к очень изменчивой обеспеченности пищей, в том числе к очень лабильному кормовому ареалу и концентрации кормов. Численность популяции этих рыб при улучшении обеспеченности пищей может быстро увеличиваться, а при ухудшении сокращаться. И. И. Николаев (1958) очень наглядно показал, что многочисленные поколения атлантическо-скандинавских сельдей, как правило, формируются на следующий год или через год после хорошего нагула маточного стада. При этом у атлантическо-скандинавских сельдей в послеличиночный период численность популяций мало сказывается на условиях нагула (Никольский и Белянина, 1959). Условия нагула этих сельдей, в том числе величина их кормового ареала, в очень значительной степени зависят от гидрологических условий года (Федоров, 1960; Шубников, 1960). У трески, нагул которой тесно связан с распространением и численностью сельди, в первую очередь ее неполовозрелых возрастных групп, условия нагула очень сильно зависят от обилия сельди и очень сильно меняются по годам, что отражается как на количестве жира в ее печени (Hjort, 1914), так и на темпе роста (Дементьева, 1967).
У азовской хамсы Engraulis encrassicholusmaeoticus Pous. наблюдается четкая многолетняя связь между жирностью родительского стада осенью и урожаем молоди на следующий год (рис. 96), т. е. условия нагула в значительной степени определяют урожай молоди (Тараненко, 1964).
|
|
|
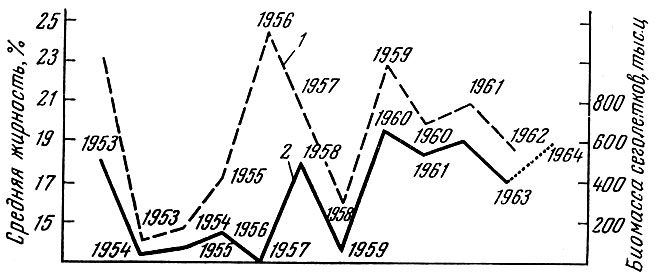
Рис. 96. Зависимость между жирностью хамсы осенью (в % к сырой массе) (1) и биомассой сеголетков на 1 сентября (в тыс. ц) (2) за 1957-1964 гг. (по Тараненко, 1964, с добавлениями)
Определенная связь существует и между условиями нагула пеламиды и численностью одного из основных видов ее корма - сардины. Видимо, сходная связь имеет место также между численностью других сардин и условиями нагула питающихся ими тунцов.
Рыбы, у которых не обнаруживается значительных колебаний урожайности, характеризуются более стабильными условиями нагула и более постоянной величиной кормового ареала. Особенно четко эта закономерность выражена у различных видов камбал. Сходная закономерность часто наблюдается у разных форм одного и того же вида. Обычно у полупроходных форм, у которых обеспеченность пищей и величина нагульного ареала маточного стада изменчивее, чем у жилых того же вида, наблюдаются и значительно большие флюктуации, чем у жилых форм. Подобное явление хорошо выражено у плотвы и ее полупроходных форм (вобла, тарань, аральская вобла). У полупроходных форм плотвы флюктуации выражены гораздо сильнее, чем у жилых. Сходная картина, видимо, имеет место у жилого и полупроходного леща, а также у жилого и полупроходного судака
Таким образом, изменчивость условий нагула маточного стада теснейшим образом связана с урожайностью потомства (Николаев, 1958, 1958а; Никольский, 1961). Флюктуации - это приспособления, выработавшиеся у рыб, живущих в условиях сильно изменчивой обеспеченности пищей, в том числе большой лабильности кормового ареала. Через изменение количества и качества продуцируемого потомства стадо производителей приспособительно отвечает на изменение условий нагула, в том числе на изменение величины нагульного ареала.
Если флюктуации являются приспособлением к изменению обеспеченности пищей, то, следовательно, механизм их должен быть как-то связан с изменениями хода обмена веществ у родительских особей. Чем благоприятнее условия нагула родителей, тем многочисленнее должно быть (конечно, при благоприятных условиях развития) потомство.
|
|
|
В отношении многих видов рыб установлено, что чем лучше были условия нагула, чем больше жиров отложено в теле рыбы, тем выше плодовитость и тем обычно больше жира отлагается в икре. Эта закономерность отмечена у салаки (Анохина, 1959, 1961, 1969), корюшки (Белянина, 1964), воблы (Чепракова, 1960) и некоторых других рыб. И в одном и том же стаде более жирные особи оказываются и более плодовитыми. По-видимому, поскольку длина рыбы и размеры икринки коррелируют, как это отметила у салаки Л. А. Раннак (1958а), то быстрее растущие особи, т. е. нагуливавшиеся в более благоприятных условиях, продуцируют и более крупную икру. Однако подобная связь наблюдается не только у видов со значительными флюктуациями, но и у рыб со слабо выраженными колебаниями урожайности по годам.
Среди рыб с резко выраженными флюктуациями нет видов, у которых икра содержала бы много жира. Так, у океанической сельди процент жира в икре колеблется от 0,47 до 4,84 (данные Нидгема, 1931; Кизеветтера, 1942, и лаборатории технологии ВНИРО); у трески жир составляет около 1,7-2,0% сырой массы икры.
У тех же видов рыб, у которых не наблюдается значительных флюктуации, жирность икры обычно оказывается выше. У осетровых она составляет более 20% сухого вещества, или 10-20% сырого. Некоторое исключение из этого правила представляют рыбы с пелагической икрой. У этих рыб как с выраженными флюктуациями (треска, пикша), так и без флюктуации (камбалы) количество жира в икре оказывается небольшим; причем у камбал в формировании поколения обеспеченность пищей на этапе смешанного питания, видимо, имеет меньшее значение, чем в момент перехода к донному образу жизни (на это указывал еще Гарстанг). Интересно, что и изменчивость жирности икры у видов со значительными флюктуациями, по-видимому, больше, чем у видов, у которых флюктуации не выражены. Так, например, у ходовой севрюги в устье Куры жирность икры колеблется от 12,45 до 17,06%, у осетра соответственно от 11,20 до 17,35%, а у океанической сельди от 0,4 до 4,2%, т. е. в первом случае колебания меньше чем в 2 раза, во втором случае более чем в 10 раз. На основании изложенных материалов можно сделать вывод, что флюктуации- это приспособление вида к изменениям обеспеченности пищей.
Изменения обеспеченности пищей могут зависеть от изменения количества и концентрации пищи, продолжительности сезона нагула в результате изменения нагульного ареала или изменения численности популяции. Механизм флюктуации в значительной степени связан с обеспеченностью внутренней пищей, полученной рыбой от родительского организма. Значительные флюктуации наблюдаются у видов рыб с малым количеством жира в икре и у которых (например, у сельди) особи на этапе смешанного питания не могут долго жить без внешней пищи. Этап смешанного питания обычно является тем периодом, когда у рыб с сильно выраженными флюктуациями формируется численность поколения. Поскольку запас и качество желтка в икре связаны с условиями нагула родительского стада за предыдущий год или предыдущие годы, то естественно, обеспеченность пищей родительского стада в значительной степени определяет урожай следующего года. Большое значение имеет, конечно, и увеличение плодовитости родительского стада при благоприятных условиях нагула. Однако рассматривать связь плодовитости стада и урожая молоди без учета качества половых продуктов нельзя.
|
|
|
Если недостаточная обеспеченность пищей на этапе смешанного питания может непосредственно вызывать гибель рыб и тем самым влиять на величину поколения, то на последующих этапах, хотя обеспеченность пищей и продолжает играть очень большую, часто ведущую роль в формировании половозрелого стада, но непосредственная гибель от голода становится исключительно редким явлением. На послеличиночных этапах развития ухудшение обеспеченности пищей вызывает снижение активности, замедление роста, усиление выедания хищниками, увеличение заболеваний, зараженности паразитами. Это экспериментально подтверждено В. С. Ивлевым (1955) на примере гибели воблы, больной Nascus cuticola, а также выедания голодавшей молоди воблы щукой и личинок окуня личинками плавунца.
Более истощенные особи хуже переносят зимовку. В связи с этим в прудовом рыбоводстве на зимовку сажают молодь определенной кондиции.
Если кормовые условия после прохождения этапа смешанного питания оказываются благоприятными, то все особи, слагающие поколение данного года рождения, растут более или менее сходно. Изменчивость роста у них небольшая. Поскольку же у рыб время полового созревания связано с достижением определенных размеров, то растущее в благоприятных условиях обеспеченности пищей поколение созревает в основной массе в одном возрасте. Вступление такого дружно созревшего мощного поколения в нерестовую популяцию резко увеличивает ее численность. Многие рыбы со значительными флюктуациями численности обладают очень большим и изменчивым в результате изменений гидрологических условий нагульным ареалом и часто совершают длительные миграции от мест размножения к местам нагула. У мощных поколений рыб, например трески Gadus morhua L. и сельди Clupea harengush.,миграции обычно протяженнее и нагульный ареал обширнее, чем у менее мощных поколений(Марти, 1961). То же самое наблюдается у сардин, скумбрии и ряда других морских стайных рыб.
Естественна, что величина нагульного ареала определяется не только численностью стада рыбы, но и наличием подходящих условий для нагула. Так, например, похолодание в Северной Атлантике приводит к сокращению северной части нагульного ареала трески вне зависимости от численности ее стада, что сказывается на ее обеспеченности пищей и темпе роста. С другой стороны, известны случаи, когда и при наличии благоприятных условий для питания нагульный ареал полностью не осваивается треской из-за сокращения численности ее стада. То же известно и в отношении многих других рыб, таких, как сельдь, сардина, пеламида и другие, т. е. главным образом тех, у которых имеются значительные флюктуации. Мной было показано, что миграции проходных рыб являются приспособлением, позволяющим путем расширения нагульного ареала и освоения для нагула морских вод значительно увеличить свою численность (Никольский, 1944, 19536). В отношении сельдевых А. Н. Световидов (1953) также отмечает, что мигрирующие сельди обладают большей численностью, чем близкие виды, не совершающие далеких миграций и, следовательно, осваивающие меньший нагульный ареал. Таким образом, увеличение протяженности миграций является важным регуляторным механизмом, повышающим обеспеченность пищей популяции в годы увеличения ее численности.
|
|
|
