 |


Периодические колебания численности и биомассы стад рыб
|
|
|
|
Поскольку у рыб колебания численности популяции находятся в связи (часто прямой) с изменениями обеспеченности пищей, то естественно, что климатические колебания, связанные с солнечной активностью и другими причинами, которые имеют определенную периодику, должны как-то отражаться на колебаниях численности и биомассы рыб, обусловливая периодику колебаний численности и тем самым уловов. Периодические колебания численности и биомассы животных известны с давних времен. О периодике налетов саранчи, "мышиных напастях" и других массовых изменениях численности организмов люди знали еще в древности. Такие периодические колебания численности изучены у млекопитающих и птиц и у многих насекомых (Формозов, 1935; Andrewartha a. Birch, 1954). Андреварта и Берч указывают как наиболее обычный период колебаний численности наземных позвоночных 9-10 лет. Эта периодика относится к лисе, рыси, зайцам, ястребу, куропатке, филину и многим другим. Довольно обычна для наземных позвоночных также периодика в 4 года (лемминг, песец, полевки, канюк, сорокопут и др.).
В отношении рыб на этом вопросе впервые детально остановились Хелланд-Хансен и Нансен (Helland-Hansen a. Nansen, 1909), которые установили, что периодические колебания уловов трески связаны с температурой Атлантического течения. Весьма подробно периодику колебаний стад трески и сельди изучил Йорт (Hjort, 1914), отметив связь между солнечной активностью, температурой воздуха и воды, массой гонад и массой печени трески (рис. 97). Между температурой и численностью стада, т. е. величиной уловов, устанавливается связь со сдвигом на 5-6 лет; это связывается с возрастом вступления рыб в промысел.

Рис. 97. Количество тресковой икры и жира печени (в гл) по данным уловов из района Лофотен за 1859-1907 гг. и Финмаркена за 1866-1906 гг. (в млн. шт.) (по Hjort, 1914): 1 - количество икры (из 1000 рыб); 2 - количество печеночного жира (в гл с 1000 рыб); 3 - уловы в районе Лофотен; 4 - уловы в Финмаркене; 5 - средние температуры воздуха с 1 ноября по 30 апреля у маяка Она
|
|
|
Йорт (1914) показал четкую связь между интенсивностью пятнообра-зования на солнце и массой печени у трески (за период 1880-1911 гг.). Эта же намеченная Йортом связь прослежена другими учеными (Маслов, 1944, 1957; Rollefsen, 1949, 1954; Ижевский, 1981, и мн. др.).
Как видно из табл. 100, которая отнюдь не может претендовать на полноту сведений, периодические колебания численности отмечены у очень многих видов рыб морских и континентальных водоемов. А. Н. Державин (1922) установил, что изменения численности стада куринской севрюги, как и ряда других рыб Каспия - осетра, воблы, жереха-хашама, судака, характеризуются примерно 11-летней периодичностью. Наблюдаются, правда, менее ясно выраженные колебания численности севрюги с периодом несколько больше 30 лет.

Таблица 100. Периодика колебаний численности рыб
У основных рыб Волго-Каспийского района (Чугунов, 1928) также намечается ясная периодичность уловов (рис. 98). Эта периодичность уловов основных промысловых рыб Северного Каспия отмечается и в последующие годы (Танасййчук, 1959) (рис. 99).

Рис. 98. Колебания уловов рыбы в Волго-Каспийском районе (по Чугунову, 1928): 1 - судак; 2 - сельдь; 3 - вобла; 4 - общий улов всех видов; 5 - колебания количества солнечных пятен, %
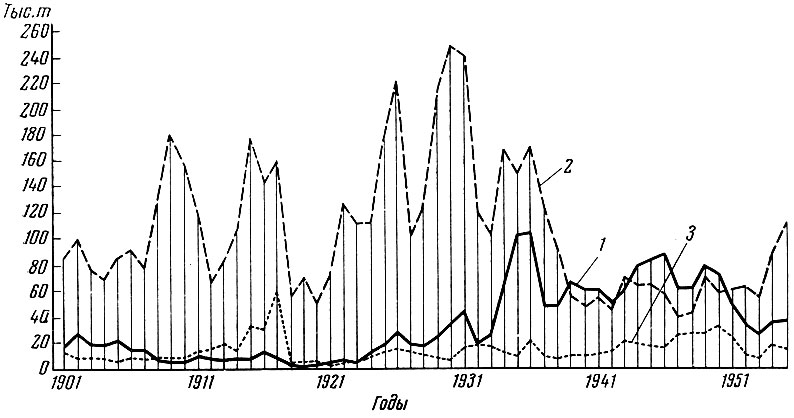
Рис. 99. Колебания уловов леща, воблы и сазана в Северном Каспии за 1901-1954 гг. (по Танасийчуку, 1959): 1 - лещ; 2 - вобла; 3 - сазан
В отношении основных промысловых рыб Северного Каспия - леща, воблы, сазана, судака, сельди, таким образом, намечается следующая схема связи: интенсивность солнечной радиации - сток Волги - урожай поколения.
Уменьшение стока связано с ухудшением условий нагула как взрослых рыб, так и молоди. Естественно, что каждый вид по-своему согласно своей специфике реагирует на эти планетарные явления, но у всех эта реакция имеет место.
|
|
|
Четкая периодика колебаний численности наблюдается и у рыб Аральского моря (Никольский, 1944; Никольский и Фортунатов, 1950; Фортунатов, Курбатова и Райская, 1950; Романычева, 1958, и др.). Большие уловы леща в 1931 г. были после максимально высокого уровня моря в 1925 г. (в уловах преобладали 6-летки); многоводный 1934 г. обеспечил высокие уловы в 1938-1939 гг. (рис. 100); наконец, высокий улов 1941 г. был получен в результате высокого урожая молоди в 1936 г. Особо благоприятные условия создаются для леща, когда высокий уровень Арала совпадает с большим стоком рек. В годы с низким уровнем Арала условия воспроизводства, особенно леща, ухудшаются и потому, что лещ во время нереста концентрируется на незначительных нерестовых площадях, где он легко поддается облову, вследствие чего неурожайность маловодных лет усиливается (Романычева, 1958). После 40-х годов в южной части Арала произошел некоторый сдвиг периодики, связанный с изменением строения дельты Амударьи.

Рис. 100. Уловы леща на юге Арала в сопоставлении со стоком Амударьи и уровнем моря (по Романычевой, 1958): 2 - уловы леща; 2 - годовой сток Амударьи; 3 - горизонт моря, абсолютные отметки. Шкала уловов смещена на 5 лет (средний возраст леща в уловах)
Периодические колебания уловов, связанные с изменением режима стока и уровня моря, удается установить и у сазана, второй основной промысловой рыбы Арала. Интересно, что в большинстве случаев (особенно четко это видно на примере леща) максимумы уловов в Арале приходятся на минимум уловов в Северном Каспии. Так, в 1934-1936 гг. в Северном Каспии уловы леща были очень высокие, а в Арале - близкие к минимуму. В 1946 г. на Арале уловы упали, а на Каспии поднялись. Максимальный (за все годы) улов на Арале был в 1939 г., на Каспии же в 1939 г. он был сравнительно невысоким.
Такая же разница в колебаниях численности отмечается и у аральской и каспийской воблы. Максимальные уловы аральской воблы были в 1932, 1937, 1942 и 1947 гг., а каспийской - в 1930, 1934-1936, 1943 и 1949 гг. Такое несовпадение максимумов и минимумов уловов одних и тех же видов в Северном Каспии и в Арале связано, как мне представляется, с тем, что усиление поступления солнечного тепла приводит к сокращению стока рек в Северном Каспии и, наоборот, к увеличению в Арал, сток которого формируется главным образом ледниковым питанием. Искусственное изменение режима стока наших южных рек путем его зарегулирования привело к изменениям, сходным с вызывавшимися космическими причинами.
|
|
|
Для леща, судака и некоторых других рыб озер Северо-Запада СССР П. В. Тюрин (1967) выявил периодичность колебаний урожайности в 11 и 16-17 лет. Он связывает повышение урожайности леща и судака с совпадением периодов потепления, с периодами высокой влажности, определяющей высокий уровень воды в озерах весной. Для байкальского омуля П. В. Тюриным (1969) выявлен ряд периодов различной длительности, связанных с общеклиматическими колебаниями и в первую очередь с состоянием уровня Байкала.
Четкая периодика в 8-11 лет выражена у атлантического лосося Salmo salar L. (Берг, 1935). Эти периоды отмечаются в пределах всего его ареала и на севере СССР, и в Норвегии, и в Канаде. Л. С. Берг на основе совпадения периодики колебаний численности лосося и пушных зверей приходит к выводу, что причины, определяющие численность поколений, действуют в речной период жизни. Иногда максимумы и минимумы уловов в разных частях ареала могут сдвигаться на год, но характер периодики выражен всюду очень четко (рис. 101).

Рис. 101. Уловы лосося по годам (по Бергу, 1935): 1 - Приморская область Канады; 2 - Норвегия; 3 - бассейн Баренцева и Белого морей
Четкая периодика уловов, связанная с климатическими изменениями, выражена у дальневосточных лососей (Бирман, 1953).
Видимо, несколько иного происхождения двухгодичная периодика у горбуши. Как правильно считают В. В. Васнецов (in litt.), А. Г. Кагановский (1949) и Ю. Е. Лапин (1971), двухгодичная периодичность численности свойственна горбуше как виду и является ее приспособлением, выработавшимся в связи с обеспеченностью пищей в морской период жизни. Как мне удалось показать (Никольский, 1953а), разница между численностью мощного и слабого поколений у горбуши тем сильнее, чем больше общая суммарная численность поколений двух смежных лет. При снижении общей численности стада сглаживается и разница в мощности поколений четного и нечетного годов. Если же рассматривать динамику численности горбуши, суммированную по двухлетиям, то удается выявить периодичность колебаний численности, как и у других дальневосточных лососей. Ю. Е. Лапин (1971), анализируя механизмы динамики численности горбуши, правильно подметил, что у горбуши поколение не всегда, видимо, состоит только из особей в возрасте 1+, но иногда включает некоторое количество трехлеток. Относительная численность трехлеток оказывается различной в поколениях разных годов рождения, что, возможно, связано со сменой годов высокой и низкой численности.
|
|
|
У красной Oncorhynchus легка (Walb.) Дальнего озера довольно чётко выражена 5-6-летняя периодика уловов (рис. 102) (Крогиус и Крохин, 1954). У красной Курильского озера продолжительность периода несколько меньше, а именно 3-5 лет (Егорова, Крогиус, Куренков и Семко, 1961).
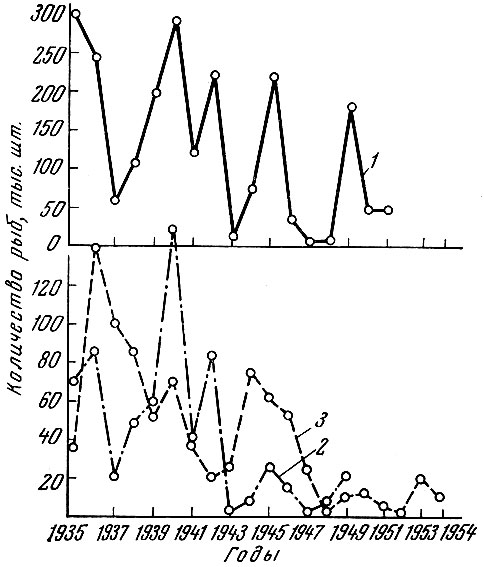
Рис. 102. Скат молоди (2), возврат на нерест (2) и пропуск производителей (3) красной в р. Дальней за 1935-1954 гг. (по Крогиус и Крохину, 1954)
У кеты и горбуши бассейна Амура И. Б. Бирман (1954, 1957, 1959) отмечает четко выраженные 10-11-летние колебания численности. Он связывает эти колебания с колебаниями стока бассейна Амура, режимом Куросио и периодами солнечной активности. Бирман отмечает, что у осенней кеты колебания урожайности выражены менее четко, чем у летней кеты и горбуши. Как было установлено мной, урожайность летней кеты Амура резко снижается в годы с низкими зимними температурами и малым количеством снега (Никольский, 1954а).
Четкая периодичность численности отмечена И. Б. Бирманом (1959) у кеты и горбуши р. Большой на Камчатке (рис. 103). Эта периодика, видимо, связана в первую очередь с деятельностью Куросио. Она оказывается у кеты сходной с периодикой, установленной А. Н. Световидовым (1952) для сахалинской сельди. Колебания численности камчатской горбуши носят противоположный характер.

Рис. 103. Динамика уловов кеты (1) и горбуши (2) (по Семко, 1954) в районе р. Большой и уловы южносахалинской сельди (3) по четырехлетиям (в % к среднему улову за все четырехлетия) (по Бирману, 1959): А - периоды похолодания моря; Б - периоды потепления (по Баталину, из Кагановского, 1949; Световидова, 1952). Пунктиром показано предполагаемое начало и окончание периодов похолодания и потепления
Таким образом, почти у всех видов рыб, о которых есть достаточно надежный материал по динамике численности и биомассы стада, удается выявить более или менее четко выраженную периодику в динамике численности, связанную с общеклиматическими изменениями. Разные исследователи устанавливают различный характер этой периодики. Так, Н. Л. Чугунов (1928) принимает периодику колебаний в 3, 6 и 11 лет. Оттестад (Ottestad, 1942) - 23/4 года, 81/2 лет, 11 лет, 171/2 лет, 23 года, 35 лет и 57 лет. Сходные периоды отмечены в росте сосны в районе Лофотен Ордингом (по Оттестаду). Оттестад отмечает, что в урожайности трески и в характере роста сосны, возможно, повинны близкие факторы. Он указывает, что урожайные или неурожайные поколения отражаются в нерестовом стаде в течение ряда лет через 6, 7, 8 лет и это несколько сглаживает величину пика. Он также указывает, что, когда пики максимумов или минимумов разных по продолжительности периодов совпадают или близки, поколение оказывается наибольшим или наименьшим.
|
|
|
Г. К. Ижевский (1961) в отношении разных рыб и разных бассейнов приводит периодичность в 2-3, 4-6, 8-10 и 18-20 лет. Он так же, как и Оттестад, отмечает возможность усиления или сглаживания явления при совпадении периодов разной продолжительности.
Если в динамике численности большинства видов рыб можно отметить отражение этой периодичности, то характер этого отражения на динамике численности у разных видов часто весьма различен. Причина этих различий заключается прежде всего в том, что рыбы разного географического происхождения (например, в Атлантике представители кельтийского фаунистического комплекса - шпрот, мерланг или арктического - мойва, сайка) по-разному реагируют на похолодание и потепление. Для налима похолодание, особенно в южной части его ареала, способствует увеличению обеспеченности пищей, а для тепловодных наоборот.
Г. К. Ижевский (1961, 1964) допускает ошибку, считая, что в пределах каждой выделяемой им области колебания численности и биомассы идут у всех видов рыб в одном направлении. Это имеет место, обычно, но и то далеко не всегда у представителей одного фаунистического комплекса, а у представителей разных комплексов колебания часто могут идти с "обратным знаком".
Потепление в Арктике и падение уровня Каспия, проявляющиеся, как это отметил Л. С. Берг (1943), более или менее синхронно, вызовут появление урожайного поколения у южнобореальных рыб Северной Атлантики и неурожайного у северокаспийского леща. Поколение же аральского леща окажется многочисленным, так как в Арале увеличится сток вследствие таяния ледников. Таким образом, и у одного вида рыб в разных частях ареала одни и те же общие климатические изменения могут вызывать разную реакцию. Г, К. Ижевский (1961), в частности, неправ, относя к одной зоне Северный Каспий и Арал, где у представителей основного Понто-Каспийского фаунистического комплекса колебания численности и биомассы обычно носят почти зеркальный характер.
Мощное поколение в последующие годы своей жизни далеко не всегда оказывается в благоприятных условиях обеспеченности пищей. Поэтому у большинства рыб часто имеется не прямая корреляция (как считал Г. К. Ижевский) между темпом роста и численностью, а обратная. Обычно с улучшением условий и появлением первых мощных поколений эти поколения оказываются и в относительно благоприятных условиях обеспеченности пищей. Однако очень скоро происходит снижение обеспеченности пищей и, при окончании благоприятного климатического цикла наступает ухудшение условий воспроизводства. У этих малочисленных поколений часто наблюдается и замедленный темп роста. Рыбы с коротким жизненным циклом реагируют на эти климатические изменения быстрее, чем рыбы с длительным жизненным циклом и более поздним достижением половозрелости.
Существенные сдвиги в периодике колебаний численности и уловов связаны со структурой популяции и в первую очередь с возрастом полового созревания. Естественно, что поздно созревающие рыбы дадут увеличение уловов (по которым мы обычно судим о динамике популяции) позднее, чем рыбы с коротким жизненным циклом. Рыбы, созревающие в течение ряда лет, дадут более сглаженную картину, чем созревающие дружнее. Периодические колебания численности и биомассы популяций - эти "волны жизни", или "взрывы жизни", как их назвал В. И. Вернадский (1927, 1934), - во много раз меняют биомассу популяций и концентрацию энергетических ресурсов в них. Эти колебания биомассы популяций сказываются как на геохимических процессах, протекающих в биосфере, так и на взаимоотношениях разных групп животных в пределах соответствующих географических зон. Такие колебания численности и биомассы промысловых видов имеют большое значение и для развития промысла (Берг, 1953).
Периодические колебания численности и биомассы стад промысловых рыб, определяемые общеклиматическими причинами, часто могут сглаживаться и меняться под влиянием действия местных факторов. Очень существенное значение при этом имеет и деятельность человека. Так, например, некоторое смещение периодики урожайности полупроходных рыб Каспийского и Азовского морей в послевоенные годы в значительной мере связано с развитием гидростроительства в бассейне этих морей.
Общей причиной периодических колебаний урожайности рыб, которые носят закономерный характер, несомненно являются периоды солнечной активности (Бирман, 1969, 1973). Но у разных видов эти общие причины преломляются по-разному, что и сказывается на разнице в характере колебаний урожайности.
Несомненно, что во многих случаях человек в состоянии уже сейчас управлять периодикой урожайности, например, путем регулирования рыболовства на местах нереста в маловодные годы, мелиоративными мероприятиями.
Зная характер периодики, уже заранее можно предусмотреть ряд мер для снижения отрицательного воздействия факторов, влияющих на урожайность.
Заключение
В трактовке общих закономерностей динамики стада сталкиваются те же противоположные взгляды, что и в трактовке закономерностей, определяющих отдельные звенья динамики популяций. Некоторые авторы еще продолжают объяснять динамику стада рыбы только как результат действия случайных внешних причин. В таких представлениях среда противопоставляется организму, ответы популяции на воздействие внешних условий трактуются как случайные, лишенные приспособительного значения. С этих же позиций отрицается связь родительское стадо - потомство. Урожай поколения связывается только с внешними причинами, приводящими к гибели ту или иную часть обычно находящегося в избытке пополнения.
Я пытался показать неправильность этой точки зрения. Как мне представляется, динамика численности и биомассы популяции - это закономерный приспособительный процесс, вытекающий из противоречивого единства внешнего и внутреннего, являющийся разрешением внутренних противоречий развития живого.
Популяция приспособительно реагирует на изменение обеспеченности пищей. Саморегуляция численности - это приспособительный процесс, это активный ответ популяции на изменение условий жизни. С представлением о приспособлениях к саморегуляции численности неразрывно связано и понимание взаимоотношений родительского стада и потомства. В известных пределах популяция может регулировать интенсивность своего воспроизводства, а не только пассивно - гибелью части особей - отвечать на изменение условий жизни.
Таким образом, основные закономерности, рассмотренные в этой главе, следующие:
1. Популяции всех видов рыб обладают системой регуляторных приспособлений, которые обеспечивают соответствие численности и биомассы популяции количеству пищи. Действие этих регуляторных приспособлений осуществляется автоматически, обычно через обмен веществ.
2. Связь численности и биомассы родительского стада и потомства носит закономерный характер. Она отражается у разных видов в определенных количественных отношениях, зависящих от уровня численности. Связь родительское стадо - потомство в известных пределах регулируется популяцией путем изменения темпа воспроизводства в связи с изменениями обеспеченности пищей. Однако, так как прямая количественная связь родители - потомство опосредуется в ряде звеньев, часто эта прямая связь нарушается и соотношение родительское стадо - потомство носит более сложный характер.
3. Колебания урожайности поколений - флюктуации - являются приспособлением к лабильной обеспеченности пищей, благодаря которому при благоприятных условиях численность рыб быстро увеличивается. Флюктуации сильно выражены у тех рыб, в желтке икринки которых жировые запасы незначительны.
4. У многих рыб, как и у других животных, довольно четко выражены периодические колебания численности и биомассы, связанные с общеклиматическими изменениями и поступлением тепла в земную атмосферу.
Эти колебания реализуются через изменения условий воспроизводства и обеспеченности пищей и увеличение тем самым выживания поколения.
Такая периодика у рыб разного географического происхождения преломляется по-разному, но у большинства видов она имеет место, часто несколько затушевываясь другими причинами.
|
|
|
