 |


Особенности пептидной связи.
|
|
|
|
Пептидная связь по своей химической природе является ковалентной и придает высокую прочность первичной структуре белковой молекулы. Являясь повторяющимся элементом полипептидной цепи и имея специфические особенности структуры, пептидная связь влияет не только на форму первичной структуры, но и на высшие уровни организации полипептидной цепи.
Большой вклад в изучение строения белковой молекулы внесли Л.Полинг и Р.Кори. Обратив внимание на то, что в молекуле белка больше всего пептидных связей, они первыми провели кропотливые рентгеноструктурные исследования этой связи. Изучили длины связей, углы под которыми располагаются атомы, направление расположения атомов относительно связи. На основании исследований были установлены следующие основные характеристики пептидной связи.

1. Четыре атома пептидной связи (С,О,N,Н) и два присоединенных
a-углеродных атома лежат в одной плоскости. Группы R и Н a-углеродных атомов лежат вне этой плоскости.
2. Атомы О и Н пептидной связи и два a-углеродных атома, а также R-группы имеют транс-ориентацию относительно пептидной связи.
3. Длина связи С–N, равная 1,32 Å, имеет промежуточное значение между длиной двойной ковалентной связи (1,21 Å) и одинарной ковалентной связи (1,47 Å). Отсюда следует, что связь С–N имеет частично ненасыщенный характер. Это создает предпосылки для осуществления по месту двойной связи таутомерных перегруппировок с образованием енольной формы,т.е. пептидная связь может существовать в кето-енольной форме.

Вращение вокруг связи –С=N– затруднено и все атомы, входящие в пептидную группу, имеют планарную транс-конфигурацию. Цис-конфигурация является энергетически менее выгодной и встречается лишь в некоторых циклических пептидах. Каждый планарный пептидный фрагмент содержит две связи с a-углеродными атомами, способными к вращению.
|
|
|
Между первичной структурой белка и его функцией у данного организма существует самая тесная связь. Для того, чтобы белок выполнял свойственную ему функцию, необходима совершенно определенная последовательность аминокислот в полипептидной цепи этого белка. Эта определенная последовательность аминокислот, качественный и количественный состав закреплен генетически (ДНК→РНК→белок). Каждый белок характеризуется определенной последовательностью аминокислот, замена хотя бы одной аминокислоты в белке приводит не только к структурным перестройкам, но и к изменениям физико-химических свойств и биологических функций. Имеющаяся первичная структура предопределяет последующие (вторичную, третичную, четвертичную) структуры. Например, в эритроцитах здоровых людей содержится белок– гемоглобин с определенной последовательностью аминокислот. Небольшая часть людей имеет врожденную аномалию структуры гемоглобина: их эритроциты содержат гемоглобин, у которого в одном положении вместо глутаминовой кислоты (заряженной, полярной) содержится аминокислота валин (гидрофобная, неполярная). Такой гемоглобин существенно отличается по физико-химическим и биологическим свойствам от нормального. Появление гидрофобной аминокислоты, приводит к возникновению «липкого» гидрофобного контакта (эритроциты плохо передвигаются в кровеносных сосудах), к изменению формы эритроцита (из двояковогнутого в серповидный), а также к ухудшению переноса кислорода и т.д. Дети, родившееся с этой аномалией, в раннем детстве погибают от серповидноклеточной анемии.
Исчерпывающие доказательства в пользу утверждения, что биологическая активность определяется аминокислотной последовательностью, были получены, после искусственного синтеза фермента рибонуклеазы (Меррифилд). Синтезированный полипептид с той же аминокислотной последовательностью, что и естественный фермент, обладал такой же ферментативной активностью.
|
|
|
Исследования последних десятилетий показали, что первичная структура закреплена генетически, т.е. последовательность аминокислот в полипептидной цепи определяется генетическим кодом ДНК, и, в свою очередь определяет вторичную, третичную и четвертичную структуры белковой молекулы и ее общую конформацию. Первым белком, у которого была установлена первичная структура, был белковый гормон инсулин (содержит 51 аминокислоту). Это было сделано в 1953 г. Фредериком Сэнгером. К настоящему времени расшифрована первичная структура более десяти тысяч белков, но это очень небольшое количество, если учесть, что в природе белков около 1012. В результате свободного вращения полипептидные цепи способны скручиваться (складываться) в различные структуры.
Вторичная структура. Под вторичной структурой белковой молекулы понимают способ укладки полипептидной цепи в пространстве. Вторичная структура белковой молекулы образуется в результате того или иного вида свободного вращения вокруг связей, соединяющих a-углеродные атомы в полипептидной цепи.В результате этого свободного вращения полипептидные цепи способны скручиваться(складываться) в пространстве в различные структуры.
В природных полипептидных цепях обнаружены три основных типа структуры:
- a-спираль;
- β-структура(складчатый лист);
- статистический клубок.
Наиболее вероятным типом строения глобулярных белков принято считать α-спираль Закручивание происходит по часовой стрелке (правый ход спирали), что обусловлено L-аминокислотным составом природных белков. Движущей силой в возникновении α-спирали является способность аминокислот к образованию водородных связей. R-группы аминокислот направлены наружу от центральной оси a-спирали. диполи >С=О и >N–Н соседних пептидных связей ориентированы оптимальным образом для дипольного взаимодействия, образуя вследствие этого обширную систему внутримолекулярных кооперативных водородных связей, стабилизирующих a-спираль.
|
|
|

Шаг спирали (один полный виток) 5,4Å включает 3,6 аминокислотных остатка.

Рисунок 2 – Структура и параметры a-спирали белка
Для каждого белка характерна определенная степень спирализации его полипептидной цепи
Спиральную структуру могут нарушить два фактора:
1) в наличие в цепи остатка пролина, циклическая структура которого вносит излом в полипептидную цепь – нет группы –NН2, поэтому невозможно образования внутрицепочечной водородной связи;
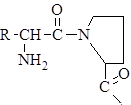
2) если в полипептидной цепи подряд расположено много остатков аминокислот, имеющих положительный заряд (лизин, аргинин) или отрицательный заряд (глутаминовой, аспарагиновой кислот), в этом случае сильное взаимное отталкивание одноименнозаряженных групп (–СОО– или –NН3+) значительно превосходит стабилизирующее влияние водородных связей в a-спирали.
Другой тип конфигурации полипептидных цепей, обнаруженный в белках волос, шелка, мышц и в других фибриллярных белках, получил название β-структуры или складчатого листа. Структура типа складчатого листа также стабилизирована водородными связями между теми же диполями –NН......О=С<. Однако в этом случае возникает совершенно иная структура, при которой остов полипептидной цепи вытянут таким образом, что имеет зигзагообразную структуру. Складчатые участки полипептидной цепи проявляют кооперативные свойства, т.е. стремятся расположиться рядом в белковой молекуле, и формируют параллельные

одинаковонаправленные полипептидные цепи или антипараллельные,
которые укрепляются благодаря водородным связям между этими цепями. Такие структуры называются b-складчатые листы (рисунок 2).

Рисунок 3 – b-структура полипептидных цепей
a-Спираль и складчатые листы – это упорядоченные структуры, в них имеется регулярная укладка аминокислотных остатков в пространстве. Некоторые участки полипептидной цепи не имеют какой-либо правильной периодической пространственной организации, их обозначают как беспорядочный или статистический клубок.
|
|
|
Все эти структуры возникают самопроизвольно и автоматически вследствие того, что данный полипептид имеет определенную аминокислотную последовательность, которая предопределена генетически. a-спирали и b-структуры обуславливают определенную способность белков к выполнению специфических биологических функций. Так, a-спиральная структура (a-кератин) хорошо приспособлена к тому, чтобы образовывать наружные защитные структуры-перья, волосы, рога, копыта. b-структура способствует образованию гибких и нерастяжимых нитей шелка и паутины, а конформация белка коллагена обеспечивает высокую прочность на разрыв, необходимую для сухожилий. Наличие только a-спиралей или b-структур характерно – для нитевидных (фибрилярных белков). В составе глобулярных (шаровидных) белков содержание a-спиралей и b-структур и бесструктурных участков сильно варьируется. Например: инсулин спирализованна 60%, фермент рибонуклеаза – 57%, белок куриного яйца лизоцим – на 40%.
Третичная структура. Под третичнойструктурой понимают способ укладки полипептидной цепи в пространстве в определенном объеме.
Третичная структура белков образуется путем дополнительного складывания пептидной цепи содержащей a-спираль, b-структуры и участки беспорядочного клубка. Третичная структура белка формируется совершенно автоматически, самопроизвольно и полностью предопределяется первичной структурой и имеет непосредственное отношение к форме белковой молекулы, которая может быть различной: от шарообразной до нитевидной. Форма белковой молекулы характеризуется таким показателем, как степень асимметрии (отношение длинной оси к короткой). У фибриллярных или нитевидных белков степень асимметрии больше 80. При степени асимметрии меньше 80 белки относятся к глобулярным. Большинство из них имеет степень асимметрии 3-5, т.е. третичная структура характеризуется достаточно плотной упаковкой полипептидной цепи, приближающейся по форме к шару.
При формировании глобулярных белков неполярные гидрофобные радикалы аминокислот группируются внутри белковой молекулы, в то время как полярные радикалы ориентируются в сторону воды. В какой-то момент возникает термодинамически наиболее выгодная стабильная конформация молекулы – глобула. В такой форме белковая молекула характеризуется минимальной свободной энергией. На конформацию возникшей глобулы оказывают влияние такие факторы как рН раствора, ионная сила раствора, а также взаимодействие белковых молекул с другими веществами.
Основной движущей силой в возникновении трехмерной структуры, является взаимодействие радикалов аминокислот с молекулами воды.
|
|
|
Фибриллярные белки. При формировании третичной структуры не образуют глобул – их полипептидные цепи не сворачиваются, а остаются вытянутыми в виде линейных цепей, группируясь в волокна- фибриллы.


Рисунок – Строение коллагеновой фибриллы (фрагмент).
В последнее время появились доказательства, что процесс формирования третичной структуры не является автоматическим, а регулируется и контролируется специальными молекулярными механизмами. В этом процессе задействованы специфические белки – шапероны. Основными функциями их являются способность предотвращать образование из полипептидной цепи неспецифических (хаотичных) беспорядочных клубков, и обеспечение доставки (транспорта) их к субклеточным мишеням, создавая условия для завершения свертывания белковой молекулы.
Стабилизация третичной структуры обеспечивается благодаря нековалентным взаимодействиям между атомными группировками боковых радикалов.

Рисунок 4- Типы связей, стабилизирующих третичную структуру белка
а) электростатические силы притяжения между радикалами, несущими противоположно заряженные ионные группы (ион-ионные взаимодействия), например отрицательно заряженная карбоксильная группа (– СОО–) аспарагиновой кислоты и (NН3+) положительно заряженной e-аминогруппой остатка лизина.
б) водородные связи между функциональными группами боковых радикалов. Например, между ОН- группой тирозина и карбоксильным кислородом аспарагиновой кислоты
в) гидрофобные взаимодействия обусловлены силами Ван-дер-Ваальса между неполярными радикалами аминокислот. (Например, группами
–СН3 – аланина, валина и т.д..
г) диполь-дипольные взаимодействия
д) дисульфидные связи (–S–S–) между остатками цистеина. Эта связь очень прочная и присутствует не во всех белках. Важную роль эта связь играет в белковых веществах зерна и муки, т.к. оказывает влияние на качество клейковины, структурно-механические свойства теста и соответственно на качество готовой продукции – хлеба и т.д.
Белковая глобула не является абсолютно жесткой структурой: в известных приделах возможны обратимые перемещения частей пептидной цепи относительно друг друга с разрывом небольшого количества слабых связей и образования новых. Молекула как бы дышит, пульсирует в разных своих частях. Эти пульсации не нарушают основного плана конформации молекулы, подобно тому, как тепловые колебания атомов в кристалле не изменяют структуру кристалла, если температура не настолько велика, что наступает плавление.
Только после приобретения белковой молекулой естественной, нативной третичной структуры он проявляет свою специфическую функциональную активность: каталитическую, гормональную, антигенную и т.д. Именно при образовании третичной структуры происходит формирование активных центров ферментов, центров ответственных за встраивание белка в мультиферментный комплекс, центров, ответственных за самосборку надмолекуляных структур. Поэтому любые воздействия (термические, физические, механические, химические), приводящие к разрушению этой нативной конформации белка (разрыв связей), сопровождается частичной или полной потерей белком его биологических свойств.
Изучение полных химических структур некоторых белков показало, что в их третичной структуре выявляются зоны, где сконцентрированы гидрофобные радикалы аминокислот, и полипептидная цепь фактически обматывается вокруг гидрофобного ядра. Более того, в ряде случаев в белковой молекуле обособляются два и даже три гидрофобных ядра, в результате возникает 2-х или 3-х ядерная структура. Такой тип строения молекулы характерен для многих белков, обладающих каталитической функцией (рибонуклеаза, лизоцим и т.д.). Обособленная часть или область молекулы белка обладающая в определенной степени структурной и функциональной автономией называется доменом. У ряда ферментов, например, обособленны субстрат-связывающие и кофермент связывающие домены.
В биологическом отношении фибриллярные белки играют очень важную роль, связанную с анатомией и физиологией животных. У позвоночных на долю этих белков приходится 1/3 от их общего содержания. Примером фибрилярных белков может служить белок шелка – фиброин, который состоит из нескольких антипараллельных цепей со структурой складчатого листа. Белок a-кератин содержит от 3-7 цепей. Коллаген имеет сложную структуру, в которой 3 одинаковые левовращающие цепи скручены вместе с образованием правовращающей тройной спирали. Эта тройная спираль стабилизирована многочисленными межмолекулярными водородными связями. Наличие таких аминокислот, как гидроксипролина и гидроксилизина также вносит вклад в образование водородных связей, стабилизирующих структуру тройной спирали. Все фибриллярные белки плохо растворимы или совсем нерастворимы в воде, так как в их составе содержится много аминокислот, содержащих гидрофобные, нерастворимые в воде R-группы изолейцин, фенилаланин, валин, аланин, метионин. После специальной обработки нерастворимый и неперевариваемый коллаген превращается в желатин-растворимую смесь полипептидов, который затем используют в пищевой промышленности.
Глобулярные белки. Выполняют разнообразные биологические функции. Они выполняют транспортную функцию, т.е. переносят питательные вещества, неорганические ионы, липиды и т.д. К этому же классу белков принадлежат гормоны, а также компоненты мембран и рибосом. Все ферменты тоже глобулярные белки.
Четвертичная структура. Белки содержащие две или большее число полипептидных цепей называют олигомерными белками, для них характерно наличие четвертичной структуры.

Рисунок – Схемы третичной (а) и четвертичной (б) структур белка
В олигомерных белках каждая из полипептидных цепей характеризуется своей первичной, вторичной и третичной структурой, и называется субъединицей или протомером Полипептидные цепи (протомеры) в таких белках могут быть либо одинаковыми либо разными. Олигомерные белки называют гомогенными, если их протомеры одинаковы и гетерогенными, если их протомеры различны. Например-белок гемоглобин состоит из 4-х цепей: двух -a и двух -b протомеров. Фермент a-амилаза состоит из 2-х одинаковых полипептидных цепей. Под четвертичной структурой понимают расположение полипептидных цепей (протомеров) относительно друг друга, т.е. способ их совместной укладки и упаковки. При этом протомеры взаимодействуют друг с другом не любой частью своей поверхности, а определенным участком (контактной поверхностью). Контактные поверхности имеют такое расположение атомных группировок, между которыми возникают водородные, ионные, гидрофобные связи. Кроме того, геометрия протомеров также способствует их соединению. Протомеры подходят друг к другу, как ключ к замку. Такие поверхности называются комплиментарными. Каждый протомер взаимодействует с другим во множестве точек, это приводит к тому, что соединение с другими полипептидными цепями или белками невозможно. Такие комплиментарные взаимодействия молекул лежат в основе всех биохимических процессов в организме.
ИЛЬИЧЕВА
|
|
|
