 |


Вторичная и третичная структуры ДНК
|
|
|
|
Нуклеотидный состав ДНК (независимо от источников ее выделения) имеет общие закономерности, которые известны как правила Чаргаффа (по имени ученого, сформулировавшего эти правила).
1. Число пуриновых оснований (А+Г) равно числу пиримидиновых оснований (Т+Ц), т. е. отношение пуринов к пиримидинам равно единице.
2. Число остатков аденина равно числу остатков тимина, т.е. отношение аденина к тимину равно единице (А/Т = 1,0)
Эти количественные соотношения были подтверждены исследованиями других ученых и стали важной предпосылкой при установлении трехмерной структуры ДНК и помогли понять каким образом генетическая информация кодируется в ДНК и передается от одного поколения к другому.
Базируясь на данных рентгеноструктурного анализа и правилах Чаргаффа Дж. Уотсон и Ф. Крик в 1953г. предложили следующую модель строения ДНК. Согласно этой модели, молекула ДНК состоит из двух полинуклеотидных антипараллельных цепей (5/®3/)(3/®5/) спирально право-закрученных одна относительно другой таким образом, что углеводнофосфатная цепь находится снаружи, а пуриновые и пиримидиновые основания внутри перпендикулярно центральной оси. (рисунок 10)
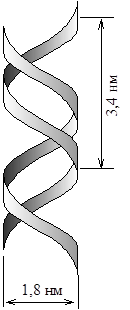
Рисунок 10 – Схематическое изображение двойной спирали ДНК
Эти две цепи соединяются между собой водородными связями, возникающими между пуриновыми и пиримидиновыми основаниями отдельных нуклеотидов, образуя специфические пары.
Тимин связан двумя водородными связями с аденином Т ::: А, цитозин тремя водородными связями с гуанином Г ::: Ц. Эти пары оснований называются комплементарными парами оснований. (рисунок 9).

Рисунок 9 – Схема образования водородных связей между комплементарными азотистыми основаниями
|
|
|
Благодаря этому нуклеотидная последовательность одной цепи полностью комплементарна последовательности другой.
Парные основания могут охватывать миллионы оснований в ДНК. Это возможно только тогда, когда полярность обоих нитей различна, т.е, когда нити имеют различное направление (различную ориентацию). Кроме того, обе нити должны быть скручены друг вокруг друга в виде двойной спирали. РНК не может образовывать из-за стерических помех, благодаря 2` - ОН групп рибозных остатков, подобную двойную спираль. Поэтому в РНК попарное соединение азотистых оснований находят только в пределах коротких участков одной и той же нити, и структура в целом менее регулярна, чем для ДНК.
Водородные связи между парами оснований – не единственный вид взаимодействий, стабилизирующих двухцепочечную структуру. Молекула ДНК – полианион, и на ее поверхности локализовано множество отрицательных зарядов, что обеспечивает стабилизацию путем электростатических взаимодействий с неорганическими противоионами, например с Mg+2,или белками, содержащими большое количество положительно заряженных боковых цепей аминокислот – гистонами. Третий стабилизирующий фактор возникает благодаря гидрофобным взаимодействиям между азотистыми основаниями, которые уложены стопкой внутри спирали. Между нитями по всей длине ДНК лежат углубления – маленькая и большая бороздки.
Так как обе нити удерживаются вместе благодаря нековалентным взаимодействиям, то двойную спираль можно разделить нагреванием (денатурацией) на одиночные нити (рисунок 11).

Рисунок 11 – разделение двойной спирали ДНК на одиночные нити
При медленном охлаждении структура двойной спирали снова восстанавливается. Денатурация ДНК играет важную роль в генной инженерии. В зависимости от рН среды, ионной силы раствора, концентрации воды и т.п. конфигурация двойной спирали может меняться. Методами рентгеноструктурного анализа доказано существование более десяти форм ДНК, которые различаются количеством пар оснований приходящихся на один виток, углом наклона оснований к вертикальной оси. Наиболее изучены А-, В-, С- и Т-формы ДНК. Предполагают, что каждая форма ДНК приспособлена для выполнения определенной биологической функции. А-форма ДНК с передачей информации от ДНК к РНК,
В-форма – с биосинтезом ДНК и С-форма с хранением, упаковкой ДНК.
|
|
|
В последние годы появились данные о возможности существования левозакрученной биспиральной молекулы ДНК-Z-формы и SBS формы ДНК, у которой полидезоксирибонуклеотидные цепи располагаются бок о бок (лесенкой, без закручивания). Такая форма ДНК обеспечивает легкое распаривание и расхождение цепей ДНК, что очень важно при биосинтезе ДНК.
ДНК обладает специфической третичной структурой. Двухцепочечная спираль ДНК на отдльных участках может подвергаться дальнейшей укладке в суперспираль. Может приобретать кольцевую форму, или свертываться в клубок. Суперскрученная структура обеспечивает экономную упаковку огромной молекулы ДНК в хромосоме. Суперспирали соединяются с белками (гистонами), упакованными в бороздах, обеспечивая тем самым стабильность третичной структуры ДНК.
Структура РНК
В клетках любых биологических объектов содержаться три основных вида РНК: рибосомальная РНК (рРНК), транспортная РНК (тРНК) и информационная или матричная РНК (мРНК). Они являются одноцепочечными молекулами различной длины, различаются по локализации, свойствам, строении, функциям. В большинстве клеток содержание РНК в 5-10 раз превышает содержание ДНК.
рРНК
Основная часть РНК клетки–70-80% приходится на долю рРНК, которая содержится в рибосомах-внутриклеточных частицах, принимающих участие в биосинтезе белка рРНК образует каркас, к которому прикрепляются белки, образуя плотноупакованный рибонуклеопротеин. Нуклеотидный состав рРНК из разных источников сходен.
мРНК
Существование матричной или информационной РНК (РНК-посредника передачи информации от ДНК в белоксинтезирующий аппарат клетки) было предсказано в 1957г., а выделена мРНК в 1962г. Содержание матричной РНК в клетке от 3% до 7% от общей суммы содержания РНК. Строение матричной РНК несколько специфично. В ее составе есть информативные участки, т.е. работающие как матрицы в процессе биосинтеза белка и неинформативные зоны. Предполагается, что неинформативные участки являются акцепторными при взаимодействии матричной РНК с рибосомой или отдельными белковыми факторами.
|
|
|
На 5`-конце молекулы РНК имеется участок, содержащий минорные нуклеотиды. Это часть так же неинформативна и называется «шапочка» или «кэп». Предполагают, что «кэп» защищает мРНК от разрушительного действия ферментов экзонуклеаз. На 3/-конце мРНК находится участок, содержащий от 50 до 400 остатков аденозинмонофосфата. Предполагают, что полиадениловый участок определяет время жизни мРНК, а так же участвует в процессе созревания и переноса м РНК из ядра в цитоплазму.
Название матричной РНК связано с функцией, которую она выполняет. Она служит матрицей, на которой синтезируется полипептидная цепь в рибосоме. Так же ее называют информационной так как она содержит информацию о том, какие аминокислоты и в какой последовательности, располагаются в белке. Эта информация представляет тройной нуклеотид, который называется кодоном. Каждый последовательно присоединенный набор из трех нуклеотидов (кодон) обеспечивает информацию для последовательного (упорядоченного) присоединения аминокислот при биосинтезе полипептида. Например, АУА-изолейцина, ГAУ-аспарагиновой кислоты и т.д. Последовательность УУУAУAГAУ (читается по три нуклеотида УУУ-AУA-ГAУ) определяет фрагмент трипептида фен-иле-асп. В клетках синтезируются тысячи различных белков, поэтому существуют тысячи матричных РНК по структуре своей комплементарных отдельным участкам ДНК.
тРНК
Содержание транспортной РНК 10% от общего содержания РНК, это самые малые по размеру молекулы РНК. Транспортная РНК не связана с клеточными структурами и находится в клетке в растворенном виде. Ее функция состоит в переносе, транспорте аминокислот к месту белкового синтеза – в рибосомы. Каждая тРНК переносит определенную аминокислоту. тРНК богаты минорными нуклеотидами.
|
|
|
Молекулы РНК, в отличие от молекул ДНК построены из одной нуклеотидной цепи, однако в этой цепи имеются комплементарные друг другу участки, которые могут взаимодействовать, образуя двойные спирали. При этом соединяются следующие нуклеотидные пары: Аденин-Урацил, Гуанин-Цитозин. Такие спирализованные участки (шпильки) обычно содержат небольшое число нуклеотидных пар и чередуются с неспаренными участками. Характерную вторичную структуру имеет транспортная РНК. Она имеет 4 спирализованные участка, и на плоскости эта структура напоминает фигуру клеверного листа. Кроме этого, имеется участок, содержащий нуклеотид комплементарный кодону матричной РНК, он называется антикодоном. С его помощью транспортная РНК прикрепляется к кодону матричной РНК. Имеется конец, который содержит остаток аденозинмонофосфата, к которому присоединяется соответствующая аминокислота (рисунок 12).

Рисунок 12 – Вторичная структура тРНК-клеверный лист
Третичная структура всех транспортных РНК схожа. Это позволяет всем им взаимодействовать с рибосомой. Незначительные отличия в пространственной структуре позволяет им взаимодействовать со специфическими ферментами тРНК синтезами, участвующими в биосинтезе белка. Третичная структура других видов РНК пока точно не установлена.
Рекомендуемая литература
1. Березов Т.Т., Коровкин Б.Ф. Биологическая химия.-М.: Медицина, 1998.-704 с.
2. Филиппович Ю.Б. Основы биохимии.-М.: Из D- во «Ангар», 1999–512 с.
3. Жеребцов Н.А., Попова Т.Н., Артюхов В.Г. Биохимия-Воронеж; Издательство ВГУ, 2002-696 с.
4. Биохимия растительного сырья. /Под. ред. В.Г. Щербакова.-М.: Колос, 1999.-376 с.
5. Ленинджер А. Биохимия.-М.: Мир, 1999 Т 1-3.
6. Основы биохимии / Под ред. А.А. Анисимова.-М.: Высшая школа, 1986.-551 с.
7. Кретович В.Л. Биохимия растений М.: Высш. школа, 1986.-503 с.
8. Химия и биохимия аминокислот и полипептидов. Методические указания для студентов технологических специальностей 49 01 01, 49 01 02, 91 01 01,48 01 02 по дисциплинам «Органическая химия», «Биологческая химия» и «Химия природных волокнообразующих полимеров». МГУП, Могилев, 2003 – 43 с.
Учебное издание
Ольга Николаевна Макасеева,
Ольга Васильевна Дудинская,
Лариса Михайловна Ткаченко,
Наталья Ивановна Ильичева
БИОЛОГИЧЕСКАЯ ХИМИЯ
Раздел «БЕЛКИ И НУКЛЕИНОВЫЕ КИСЛОТЫ»
Конспект лекций
(Учебно-методическое пособие)
Редактор А.А. Щербакова
Технический редактор Т.В. Багуцкая
Подписано в печать 14.08.2013. Формат 60×84 1/16.
Бумага офсетная. Гарнитура Таймс. Ризография.
|
|
|
Усл. печ. л. 8,5. Уч.-изд. л. 9,1.
Тираж 200 экз. Заказ 117.
Учреждение образования
«Могилевский государственный университет продовольствия».
ЛИ № 02330/630 от 31.01.2012 г.
Пр-т Шмидта, 3, 212027, Могилев.
Отпечатано в учреждении образования
«Могилевский государственный университет продовольствия».
Пр-т Шмидта, 3, 212027, Могилев.
|
|
|
