 |


Гидрофильные свойства белков. Высаливание белков
|
|
|
|
При формировании третичной структуры белков неполярные (гидрофобные) радикалы аминокислот как бы погружаются вовнутрь белковой молекулы, образуя там сухие зоны, а радикалы полярных (гидрофильных) аминокислот ориентированы в сторону воды. Происходит взаимодействие гидрофильных групп радикалов полярных аминокислот с диполями воды, в результате чего образуется водная (гидратная) оболочка белковой молекулы. (см. рисунок 7).
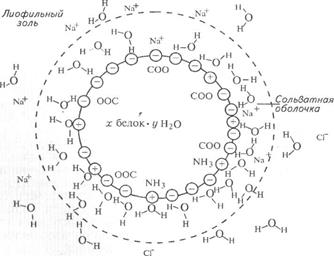
Рисунок 7 – Схема гидратной оболочки вокруг белковой молекулы
в присутствии НСl
Так, пептидная группировка, выступающая на поверхности белковой глобулы притягивает одну молекулу воды, аминогруппа –NH2 – тоже одну, а карбоксильная группа –СООН – четыре. Вблизи молекулы белка отмечается упорядоченное расположение молекул воды. По мере удаления от молекул белка расположение молекул воды носит все более беспорядочный характер. Водная оболочка является одним из факторов устойчивости белковых растворов – препятствует слипанию молекул белка и их осаждению.
При контакте сухого белка с водой он набухает, молекулы воды проникают в белковую массу, гидратируют молекулы белка, разъединяя их. Важную роль в этом процессе играют не только электростатические силы, но и силы осмоса. Дальнейшее поглощение воды приводит к растворению белка.
При определенных условиях белковые растворы образуют коллоидные системы – гели или студни, в которых растворитель и белок образуют одну внешне гомогенную массу. В гелях имеется гидратационная вода, окружающая толстым слоем коллоидные частицы белка, а так же вода, удерживаемая в капиллярных пространствах между ними. Высушенный гель, помещенный в воду, впитывает ее в очень больших количествах – набухает.
|
|
|
Гидрофильные свойства белков, т.е. их способность образовывать студни, набухать имеет большое значение в биологии и пищевых производствах. Очень подвижным студнем, является цитоплазма – полужидкое содержимое клетки, хрусталик глаза и т.д. Типичным белковым сильногидраторованным гелем является пшеничная клейковина, она содержит около 66% воды. Гидрофильные, водоудерживающие свойства продуктов необходимо так же учитывать при замораживании и размораживании мяса, овощей, фруктов, что бы сохранить, не ухудшить их качество. Гидрофильность белков зерна и муки играет большую роль при хранении и переработке зерна (кондиционирование, прорастание зерна) и хлебопечении. Студни используются в процессе приготовления различных заливных блюд.
Если у белковых глобул отнять гидратную оболочку, то частицы белка начнут слипаться, образуя ассоциации частиц белка, и выпадут в осадок. В изоэлектрической точке белки обладают наименьшей способностью связывать воду, поэтому их легче осаждать. Агрегация белковых молекул происходит и при их обезвоживании с помощью некоторых органических растворителей – например спиртом и ацетоном. Молекулы этих веществ, являясь более гидрофильными, чем молекулы белка, образуют собственные гидраты с водой, оттягивая воду от белка, лишая их водной оболочки способствуют агрегации белка.
Осаждение белков путем добавления нейтральных растворов солей высокой концентрации называется высаливанием. Осаждающая способность соли зависит от размеров катиона и аниона, а так же от величины их заряда. Катионы и анионы по высаливающей способности можно расположить в ряды, в которых эта способность убывает.
Катионы: Cs+, Rb+, K+, Na+, L i+, Ba2+, Sr2+, Co2+, Mg2+.
Анионы: SO4-2, C L -, Br, NO3-, J-, CNS-, Mg 2+.
Эти ряды называются лиотропными рядами. Высаливающее действие объясняется тем, что при высокой концентрации ионов в растворе белка, они оттягивают на себя от молекул белка поляризованные молекулы воды и тем самым лишают белок гидратной оболочки, которая препятствует осаждению белка. Метод высаливания используется для разделения и получения в очищенном виде белков и ферментов.
|
|
|
При добавлении растворов нейтральных солей (Na2SO4, (NH4)2SO4, MgSO4 и др.) небольших концентраций, растворимость белков в воде возрастает. Растворению белков, как и других веществ, способствуют те факторы, которые уменьшают взаимодействие между молекулами растворяемого вещества. Нейтральные соли в малых концентрациях увеличивают степень диссоциации ионизированных групп белковых молекул и тем самым уменьшают белок-белковое взаимодействие. Известно, что степень диссоциации электролитов (в том числе белков) прямо пропорциональна диэлектрической постоянной растворителя, которая в свою очередь пропорциональна степени поляризации молекул растворителя, их дипольному моменту. Нейтральные соли в малых концентрациях еще больше увеличивают диэлектрическую постоянную воды. В результате вода усиливает диссоциацию растворенного вещества, в частности белка. Входя между заряженными группами и ориентируясь вокруг них, диполи воды препятствуют их взаимодействию.
Растворимость белков зависит так же от рН растворителя, его состава, температуры. Минимальной растворимостью обладают белки в ИЭТ, что объясняется отсутствием электростатического отталкивания между молекулами белка.
Белки способны образовывать высококонцентрированные системы жидкось-газ-пены. В качестве традиционных пенообразователей используют белки сыворотки крови, молока, которые подвергают вначале гидролизу, а затем сушат на распылительных сушках. Устойчивость пены, в которой белок является пенообразователем зависит от его природы, концентрации и температуры.
Белки в качестве пенообразователей играют важную роль при образовании пены в пиве. В кондитерской промышленности это свойство белков используется при выработке пастилы, зефира, суфле. Структуру пены имеет хлеб и это влияет на его органолептические и структурно-механические свойства.
Благодаря гидрофильным и гидрофобным группировкам белки являются поверхностно-активными веществами и могут влиять на растворимость других веществ. Они выступают в роле эмульгаторов – веществ, стабилизирующих эмульсию, которую образуют взаимнонерастворимые жидкости (вода-жир). В организме человека в эмульгированном состоянии находятся жиры в крови и лимфе. Молоко представляет собой эмульгированные казеиногеном капельки жира в воде. Белки в качестве эмульгаторов находят применение в пищевой промышленности при производстве майонезов, кондитерских изделий, кремов, шоколада и т.п.
|
|
|
Методы выделения белков
Выделение белков из биологических тканей проводят после тщательного измельчения исследуемого материала, вплоть до разрушения клеточной структуры. Разрушают клетки механическим путем, растирая ткань с песком в ступке или в гомогенизаторе. Существуют и другие методы, например поочередное замораживание и оттаивание, обработка ультразвуком.
На всех этапах выделения и очистки белков следует учитывать их большую способность к потере природных, нативных свойств, т.е. к денатурации.
В большинстве случаев процесс разрушения клеток сопровождается выделением тепла, поэтому с целью предотвращения тепловой денатурации все операции следует проводить при пониженных температурах (около +4°С) в термостатированных холодных комнатах.
Современные методы измельчения тканей обычно сочетают с одновременной экстракцией белков из гомогенатов тканей. В качестве растворителей используют 8-10% растворы солей, различные буферные растворы, органические растворители (спирт, ацетон и т.п.), а так же неионные детергенты – вещества, разрушающие гидрофобные взаимодействия между белками и липидами и между белковыми молекулами.
После достижения полной экстракции белков, приступают к разделению-фракционированию смеси белков на индивидуальные белки. Для этого применяют различные методы: высаливание, осаждение органическими растворителями, хроматографию, электрофорез.
При выделении и очистке белков используют четыре основных вида хроматографии: адсорбционную, распределительную, ионообменную и аффинную (хроматографию по сродству). В адсорбционной хроматографии разделение компонентов смеси основано на их различной сорбируемости на твердом адсорбенте. В качестве адсорбентов используют активированный древесный уголь, оксиды алюминия или кремния. Адсорбент в виде суспензии с растворителем (чаще всего буферным раствором) вносят в колонку и равномерно утрамбовывают. Исследуемый образец в небольшом объеме растворителя вносят в колонку. Компоненты разделяемой смеси адсорбируются на адсорбенте. Затем приступают к десорбции компонентов из колонки, используя подходящие элюенты. Сбор фракций осуществляют при помощи автоматического коллектора фракций.
|
|
|
При распределительной хроматографии твердая фаза служит только опорой (основой) для стационарной жидкой фазы. Разновидностью распределительной хроматографии является хроматография на бумаге. В качестве стационарной фазы при этом служит вода, адсорбированная целлюлозными цепями фильтровальной бумаги. Образец наносят в виде капли (пятна) на одном конце бумажной полосы, этим же концом бумагу погружают в подходящую смесь органических растворителей (например, бутанол, уксусная кислота, вода в определенных соотношениях). При движении растворителя по бумаге благодаря силе капиллярности происходит разделение компонентов смеси. Проявленную хроматограмму высушивают, а местоположение каждого из разделяемых веществ определяют химическими или физико-химическими методами.
В ионообменной хроматографии в зависимости от заряда разделяемых белков используют подходящую ионообменную смолу (катионит или анионит) с функциональными группами которой обмениваются и задерживаются на колонке часть белков, в то время как другие белки беспрепятственно элюируются из колонки. Связанные с ионообменной смолой белки, отделяют, применяя более концентрированные солевые растворы или изменяя рН элюента.
Аффинная хроматография основана на принципе избирательного взаимодействия белков (или других молекул) с закрепленными на носителе специфическими веществами – лигандами, которыми могут быть субстраты или коферменты (если выделяется какой-либо фермент) и т.д. Благодаря высокой специфичности белков к иммобилизованному (закрепленному) лиганду, к нему присоединяется только один какой-либо белок из смеси. Снятие с колонки этого белка осуществляется подобранными специальными элюентами.
В гель-хроматографии в качестве стационарной фазы используют гель в виде крошечных гранул, так что такую стационарную фазу можно рассматривать как молекулярные сита. Гранулы геля изготовлены из полимера со сшитой структурой, подобной ситу. В качестве такого полимерного материала используются или сшитая агароза, или декстран (полисахарид), или сшитый полиакриламид. В водной среде полимерный материал сорбирует воду и набухает, превращаясь в гелеподобные гранулы, сохраняющие пористую структуру, причем размер пор такого набухшего материала определяется степенью сшивки полимера. Нанесенные на колонку соединения (в виде раствора в подвижной фазе) начинают взаимодействовать с гранулами геля, проникая в объем гранул через поры, что замедляет движение растворенного вещества по колонке. Молекулы небольшого размера лучше задерживаться на колонке, поскольку легче проникают в объем гранул и распределяются там. Молекулы с размерами, большими, чем размеры пор, совсем не будут проникать внутрь гранул, они первыми вымываются из колонки.
|
|
|
Результаты хроматографического разделения представляются графически в виде зависимости измеряемого свойства (поглощение в УФ области и т.п.) от объема элюата, вышедшего из колонки. Пики на таком графике соответствуют выходу индивидуальных белков. Если при разделении получается только один пик, это указывает на чистоту исходного препарата, нанесенного на колонку.
Свойство белков приобретать определенной величины заряд при данном значении рН раствора нашло широкое применение для их разделения методом электрофореза. Электрофорез основан на передвижении заряженной частицы в электрическом поле. Движение ее происходит в жидкой среде, которая удерживается инертным твердым носителем, например, полоской бумаги, гелевой пленкой из крахмала, полиакриламида, декстрона и т.д.
При постоянном напряжении движение заряженной молекулы белка определяется отношением заряда к ее размеру:
| m =¦ (Q/r), | 4.4.1 |
где Q-суммарный заряд белковой молекулы;
r-радиус молекулы.
С увеличением этого отношения подвижность молекулы растет. Так как каждый белок имеет свою определенную величину Q/r, скорость перемещения различных белков в электрическом поле будет различной. Электрофорез используется для разделения белков и определения их молекулярных масс.
Применение в определенной последовательности выше перечисленных методов позволяет получить белок в очищенном состоянии, не лишенный, однако, некоторых примесей солей. Для полного очищения белков от низкомолекулярных примесей используются методы диализа, кристаллизации, гельхроматографии и ультрафильтрации.
Является ли полученный белковый препарат индивидуальным белком или смесью имеет важное значение. Всегда можно ожидать, что в составе изолированного белка есть примесь других белков; это может привести к неправильным выводам о свойствах исследуемого белка. Поэтому большое внимание уделяется оценке гомогенности – однородности белков. Критерием чистоты белков служат следующие показатели: получение белка в кристаллическом состоянии; дальнейшая неразделяемость при электрофорезе и ультрацентрифугировании; независимость растворимости от количества твердой фазы; постоянство аминокислотного состава; определенный молекулярный вес; для многих белков – постоянство специфических биологических свойств (ферментативная активность, гормональная активность и т.д.)
Классификация белков
Белки в зависимости от химического строения делят на простые (протеины) и сложные (протеиды). Простые белки при гидролизе распадаются только на аминокислоты. При гидролизе сложных белков наряду с аминокислотами образуется вещество небелковой природы – простетическая группа. Классификация простых белков (протеинов) основана на их растворимости.
Альбумины – водорастворимые белки с высокой гидрофильностью, выпадают в осадок при 100%-ом насыщении сульфатом аммония. К этим белкам относятся белок куриного яйца, белки зародыша семян злаковых и бобовых культур. Альбумин пшеницы называют лейкозин, гороха – легумелин. Альбумины содержат все незаменимые аминокислоты.
Глобулины – растворяются в солевых растворах, чаще всего для извлечения глобулинов используют 2 –10%-ый раствор хлорида натрия. Они осаждаются 50%-ым раствором сульфата аммония. Белки семян бобовых и масличных культур в основном представлены глобулинами; легумин – гороха и чечевицы, фазеолин – фасоли; глицин – соевых бобов. Многие альбумины и глобулины обладают ферментативным действием.
Проламины. Эта группа белков характерна исключительно для семян злаков. Эти белки растворяются в 60-80%-ом растворе этилового спирта. Эти белки содержат значительные количества пролина и глютаминовой кислот. Лизина они не содержат или содержат его в следовых количествах. Хорошо изучены проламины пшеницы – глиадины, ячменя – гордеин, кукурузы – зеин. Проламины – это комплексы белков различающиеся по составу и молекулярной массе.
Глютелины находятся, как правило, с проламинами. Растворяются они в щелочах (чаще 0,2%-ым NaOH). Глютелины не однородные белки, а смеси разных белков со сходными свойствами. Наиболее исследованы глютелин пшеницы, орезенин риса.
Глютенин и глиадин пшеницы образуют комплекс, который называют клейковиной. Клейковина муки влияет на структурно-механические свойства теста, а, следовательно на качество хлеба.
Протамины – самые низкомолекулярные белки. Встречаются эти белки в молоках рыб. На 2/3 эти белки состоят из аргинина, поэтому имеют основной характер. Протамины не содержат серы.
Гистоны – содержаться в хромосомах клеточных ядер, они участвуют в стабилизации пространственной структуры ДНК. Гистоны на 20-30% состоят из основных аминокислот. Из растворов их осаждают аммиаком.
Протеноиды – подгруппа нерастворимых фибриллярных белков животного происхождения. К ним относятся фиброин – шелка, кератин – волос, рогов, перьев, сухожилий и связок. Характерная особенность протеноидов – высокое содержание в них серы. Эти белки не гидролизуются пищеварительными ферментами.
Протеиды – сложные белки, в которых белок связан с веществом небелковой природы – простетической группой. В зависимости от химической природы простетической группы их делят на липопротеины, гликопротеины, хромопротеины, нуклеопротеины, фосфопротеины, металлопротеины.
В липопротеинах простетическая группа представлена каким-либо липидом. Эти белки входят в состав клеточных мембран, участвуют в структурной организации миелиновых оболочек, нервной ткани, хлоропластов и т.д., а так же присутствуют в свободном состоянии.
Гликопротеины – в качестве простетической группы содержат углеводный компонент. Это могут быть глюкоза, манноза, N-ацетилглюкозамин, L -фукоза и т.д. Эти белки выполняют специфические функции: обеспечивают клеточную адгезию, молекулярное и клеточное узнавание, антигенную активность опухолевых клеток, оказывают защитное, гормональное, и антивирусное действие.
Типичные представители гликопротеинов – белки, входящие в состав слюны, а так же некоторых растительных слизей. Растительные гликопротеины называют еще лектинами. Некоторые лектины оказывают антипитательное действие – они нарушают процессы всасывания питательных веществ. Антипитательное действие лектинов фасоли твердо установлено.
Хромомпротеины – состоят из простого белка и связанного с ним окрашеного небелкового компонента. Среди хромопротеинов различают гемопротеины, содержащие в качестве простетической группы железо- или магнийпорфирины; флавопротеины – содержащие производные изоаллоксазина. Хромопротеины участвуют в таких функциональных процессах жизнедеятельности, как транспорт кислорода и углекислого газа, фотосинтез, окислительно-восстановительные реакции, свето- и цветовосприятие и т.д. К группе хромопротеинов относятся гемоглобин и его производные, хлорофилсодержащие белки, такие ферменты как каталаза и пероксидаза, сукцинатдегидрогеназа, все белки цитохромы.
Нуклеопротеины – одна из самых важных групп белков, состоящая из простых белков связанных с нуклеиновыми кислотами. Эти белки играют первостепенную роль в хранении и передаче генетической информации и биосинтезе белка и содержаться в основном в ядрах клеток. Дезоксирибонуклеопротеины содержат дезоксирибонуклеиновую кислоту (ДНК). Рибонуклеопротеины содержат рибонуклеиновую кислоту (РНК)
Фосфопротеины – эти белки содержат органически связанный, лабильный фосфат, абсолютно необходимый для выполнения клеткой ряда биологических функций. Кроме того, они являются ценным источником энергетического и пластического материала в процессе роста и развития зародышей и молодого растущего организма. Наиболее изучены фосфопротеины – казеин молока, вителлин яичного желтка, ихтулин икры рыб.
Металлопротеины наряду с белком содержат ионы какого-либо металла или нескольких металлов. Металлопротеины выполняют различные функции. Например, белок трансферрин (содержит железо) служит физиологическим переносчиком железа в организме. Другие металлопротеины являются биологическими катализаторами-ферментами – амилазы (содержат Са2+) гидролизуют крахмал, карбоангидроза (Zn2+) расщепляет угольную кислоту, аскорбатоксидаза (Cu2+) разрушает витамин С и т.д.
НУКЛЕИНОВЫЕ КИСЛОТЫ
Нуклеиновые кислоты были открыты в 1868г. швейцарским врачом
Ф.Мишером. Биологическая функция этого вещества оставалась неизвестной еще в течение почти столетия, и только в 40-х годах прошлого века Эвери, Маклеод и Маккарти установили, что нуклеиновые кислоты, отвечают за хранение, репликацию (воспроизведение), транскрипцию (передачу) и трансляцию (воспроизведение на белок) генетической (наследственной) информации. Нуклеиновые кислоты определяют вид, форму, химический состав и функции живой клетки и всего организма в целом.
В 1953 г. Уотсон и Крик сообщили о расшифровке молекулярной структуры ДНК. В каждом живом организме присутствуют два типа нуклеиновых кислот: рибонуклеиновая кислота (РНК) и дезоксирибонуклеиновая кислота (ДНК). В то же время вирусы содержат только один какой-нибудь тип нуклеиновых кислот: РНК, ДНК.
Нуклеиновые кислоты – это высокомолекулярные соединения, размер которых сильно варьирует. Молярная масса транспортной РНК составляет 25000, тогда как отдельные молекулы ДНК обладают массой от 1 000 000 до 1 000 000 000.
Количественное содержание ДНК в клетках одного и того же организма постоянно и исчисляется несколькими пикограммами, однако в клетках разных видов живых организмов имеются существенные количественные различия в содержании ДНК. ДНК преимущественно сосредоточено в ядре, митохондриях и хлоропластах. РНК большей частью содержится в цитоплазме клеток. Содержание РНК, как правило, в 5-10 раз больше, чем ДНК. Соотношение РНК/ДНК в клетках тем выше, чем интенсивнее в них синтез белка.
Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами и при физиологических значениях рН несут высокий отрицательный заряд. В связи с этим в клетках организмов они легко взаимодействуют с различными катионами и прежде всего с основными белками, образуя нуклеопротеины.
Состав нуклеиновых кислот
Нуклеиновые кислоты при полном их гидролизе распадаются на три типа веществ – азотистые основания (пуриновые и пиримидиновые основания), сахара (пентозы) и фосфорную кислоту.
Пентозы нуклеиновых кислот представлены D- рибозой или 2- D- дезоксирибозой. Оба эти сахара содержатся в составе нуклеиновых кислот в фуранозной форме и имеют b-конфигурацию:

Нуклеиновая кислота называется рибонуклеиновой (РНК), если в ее состав входит рибоза, или дезоксирибонуклеиновой (ДНК), если в ее состав входит дезоксирибоза. Недавно установлено, что рибоза и дезоксирибоза не являются единственными углеводами, входящими в состав нуклеиновых кислот: в ряде фаговых ДНК и РНК некоторых видов раковых клеток найдена глюкоза.
Азотистые основания, которые обычно встречаются в нуклеиновых кислотах – это производные пурина аденин (А) и гуанин (G)-и производные пиримидина – цитозин (С), тимин (Т) и урацил (U). Сами пурин и пиримидин в состав нуклеиновых кислот не входят.

Строение основных азотистых оснований-компонентов нуклеиновых кислот:

Цитозин, аденин, гуанин содержатся в нуклеиновых кислотах обоих типов, урацил входит только в состав РНК, а тимин в ДНК.
Для гуанина, цитозина, тимина и урацила известна кето-енольная таутомерия, однако кетоструктуры гораздо более стабильны и доминируют при физиологических условиях.
Таутомерия

В нуклеиновых кислотах все оксосодержащие азотистые основания присутствуют в кетоформе.
В составе ДНК и РНК встречаются так называемые необычные или «минорные» азотистые основания. К ним относятся, например,
5-метилцитозин, 4-тиоурацил, дигидроурацил и др.
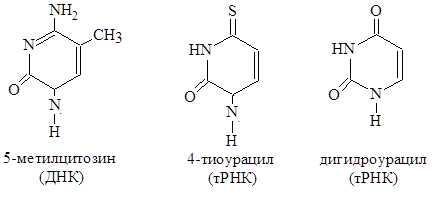
Рассмотренные пуриновые и пиримидиновые основания, а так же некоторые другие производные пурина и пиримидина, которые не входят в состав нуклеиновых кислот, часто содержатся в растениях в значительном количестве в свободном состоянии. Наиболее часто в свободном состоянии в растениях встречаются гипоксантин (6-гидроксиоксипурин), найденный в семенах горчицы и люпина. Ксантин (2,6-дигидроксиоксипурин) и аллонтоин очень широко распространены в растениях. В форме этих оснований, как и в форме амидов аминокислот, происходит запасание и транспорт азота в растениях.
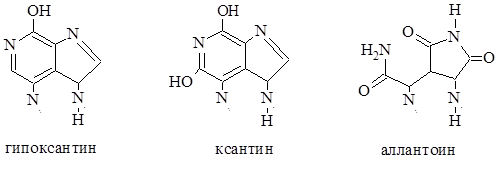
Пурины и пиримидины поглощают электромагнитную энергию в ультрафиолетовом (УФ) диапазоне, причем каждое соединение имеет характеристический спектр поглощения, однако для всех этих соединений максимум поглощения наблюдается вблизи 260 нм. Нуклеиновые кислоты так же поглощают в УФ-области. На этом свойстве основаны методы количественного определения нуклеиновых кислот.
В процессе обмена веществ у животных и растений пуриновые основания образуют такие продукты, как мочевая кислота, кофеин, теобромин, последние используются как лекарства.
Нуклеозиды
Азотистое основание с присоединенным к нему углеводным остатком, рибоза или дезоксирибоза называют нуклеозидом. В нуклеозидах ковалентная связь образована С1-атом сахара и N1- атомом пиримидина или N9- атомом пурина, такая связь называется гликозидной. Что бы избежать путаницы в нумерации, атомы углеводной части отличают штрихом. Для наиболее распространенных нуклеозидов приняты тривиальные названия: аденозин, гуанозин, уридин и цитидин. Дезоксирибонуклеозиды называются дезоксиаденозин, дезоксигуанозин, дезоксицитидин и тимидин.
Например:
Пиримидиновый Пуриновый
рибонуклеозид дезоксирибонуклеозид

Нуклеозиды являются фрагментом структуры нуклеотидов; однако многие нуклеозиды встречаются в свободном состоянии. Некоторые из них обладают лечебными свойствами. Различные микроорганизмы выделяют арабинозилцитозин и арабинозиладенин, в состав которых входит b- D- арабиноза вместо рибозы. Эти вещества используются в качестве мощных антивирусных и антигрибковых агентов и против некоторых видов рака. Механизм действия ara-А и ara-Ц основан на ингибирование биосинтеза ДНК.

Нуклеотиды
Нуклеотиды – это фосфорные эфиры нуклеозидов. В образовании связи участвует 51-углеродный атом пентозы. В зависимости от строения пентозы все нуклеотиды можно разделить на рибонуклеотиды и дезоксирибонуклеотиды.
В зависимости от числа имеющихся остатков фосфорной кислоты различают нуклеозидмонофосфаты, нуклеозиддифосфаты и нуклеозидтрифосфаты. Все эти три вида нуклеотидов постоянно присутствуют в клетках.

Рисунок 8 – моно-, ди- и трифосфонуклеотиды (51) аденозина
Названия отдельных нуклеотидов часто обозначают сокращенно большими первыми буквами названий соответствующих оснований. Ниже приведены нуклеотиды, входящие в состав нуклеиновых кислот, и даны их условные сокращенные обозначения.
Таблица 2 – Сокращенные названия отдельных нуклеотидов
| Основание | Нуклеотиды в составе РНК | Нуклеотиды в составе ДНК | Сокращенное обозначение |
| Аденин Гуанин Цитозин Урацил Тимин | Адениловая кислота Гуаниловая кислота Цитидиловая кислота Урациловая кислота Тиминовая кислота | Дезоксиадениловая кислота Дезоксигуаниловая кислота Дезоксицитидиловая кислота Дезоксиуридиловая кислота Дезокситимидиловая кислота | А Г Ц У Т |
Нуклеотиды – это сильные кислоты, так как остаток фосфорной кислоты, входящий в их состав, сильно ионизирован.
Главная функция нуклеотидов в клетке состоит в том, что они являются составными частями нуклеиновых кислот.
Все нуклеозиддифосфаты и нуклеозидтрифосфаты содержат высокоэнергетические связи (обозначены значком «~»). При гидролизе этой связи освобождается от 30 до 50 кДж/моль энергии, в то время как при гидролизе обычной сложноэфирной фосфатной связи освобождается энергия равная 8-12 кДж/моль.
Под влиянием соответствующих ферментов фосфатные группы содержащие высокоэнергетические связи, могут быть перенесены на другие вещества. Таким образом, энергия, накопившаяся в высокоэнергетических соединениях, может быть использована далее в обмене веществ. Например: АДФ и АТФ принимают участие в биосинтезе белка. Уридинтрифосфат (УТФ) и уридиндифосфат (УДФ) необходимы для действия ферментов, катализирующих превращения и синтез сахаров (ЦДФ и ЦТФ) цитидиндифосфат и цитидинтрифосфат принимают участие в биосинтезе фосфолипидов.
Циклические нуклеотиды были выделены в 1959г. Сазерлендом
(лауреат Нобелевской премии 1971г.) при изучении механизма действия некоторых гормонов при регулировании метаболизма углеводов. В циклических нуклеотидах фосфорная кислота связывает два атома кислорода пентозного остатка в одном и том же нуклеотиде. Известны три циклических нуклеотида – циклический аденозинмонофосфат (ц АМФ), циклический гуанозинмонофосфат (ц ГМФ) и циклический цитозинмонофосфат (ц ЦМФ).
Эти нуклеотиды образуются из соответствующих нуклеозидтри-фосфатов под действием ферментов аденилатциклазы и гуанилатциклазы. В биологических процессах они выступают в качестве промежуточного посредственника регуляторного действия гормонов.

Фосфатные остатки могут образовывать друг с другом кислотные ангидриды. Поэтому у нуклеотидов имеется возможность связываться друг с другом через фосфатные остатки. При этом возникают динуклеотиды с фосфоангидридной структурой.
К этой группе относятся некоторые кофакторы ферментов – НАД+ (никотинамидадениндинуклеотид), и НАДФ+ (никотинамидадениндинуклеотидфосфат), ФАД (флавинадениндинуклео-тид) и т. д.

НАД+
|
|
|
