 |


161.Транспорт газов кровью. Соединение гемоглобина с газами.
|
|
|
|
160. Газообмен в лёгких. Постоянство состава альвеолярного воздуха. Значение для газообмена в лёгких градиента парциального давления газов в альвеолярном воздухе и их напряжения в крови капилляров. Факторы, способствующие диффузии газов в легких. Закон Фика. Диффузионная способность легких.
Газообмен в лёгких:
Важнейшая функция легких – обеспечение газообмена между воздухом легочных альвеол и кровью капилляров малого круга кровообращения. Для понимания механизмов газообмена необходимо знать газовый состав этих обменивающихся между собой сред, свойства альвеолокапиллярных структур, через которые идет газообмен, и учитывать особенности легочного кровотока и вентиляции, рассмотренные выше:
Значение для газообмена в лёгких градиента парциального давления газов в альвеолярном воздухе и их напряжения в крови капилляров:

Состав атмосферного, альвеолярного (содержащегося в легочных альвеолах) и выдыхаемого воздуха представлен в табл. 2. 1.
На основе определения процентного содержания газов в альвеолярном воздухе рассчитывают их парциальное давление (Давление, которое имел бы газ, входящий в состав газовой смеси, если бы он один занимал объём, равный объёму смеси при той же температуре). При расчетах давление водяного пара в альвеолярном газе принимают равным 47 мм рт. cт.
*Например, если содержание кислорода в альвеолярном газе равно 14, 4%, а атмосферное давление – 740 мм рт. cт., то парциальное давление кислорода (рО2) составит: рО2 = (740 – 47)/100 · 14, 4 = 99, 8 мм рт. cт.
В условиях покоя парциальное давление кислорода в альвеолярном газе колеблется около 100 мм рт. cт., а парциальное давление углекислого газа – около 40 мм рт. cт. Несмотря на чередование вдоха и выдоха при спокойном дыхании состав альвеолярного газа изменяется лишь на 0, 2– 0, 4%, поддерживается относительное постоянство состава альвеолярного воздуха и газообмен между ним и кровью идет непрерывно. *
|
|
|
Постоянство состава альвеолярного воздуха поддерживается благодаря малой величине коэффициента вентиляции легких ( КВЛ ). Этот коэффициент показывает, какая часть функциональной остаточной емкости обменивается на атмосферный воздух за один дыхательный цикл. В норме КВЛ равен 0, 13–0, 17 (т. е. при спокойном вдохе обменивается приблизительно 1/7 часть ФОЕ ).
Состав альвеолярного газа по содержанию кислорода и углекислого газа на 5–6% отличается от атмосферного. Поскольку коэффициент вентиляции различных областей легких может отличаться, состав альвеолярного газа может иметь разную величину не только в отдаленных, но и в соседних участках легкого. Это зависит от диаметра и проходимости бронхов, выработки сурфактанта и растяжимости легких, положения тела и степени наполнения кровью легочных сосудов, скорости и соотношения длительностей вдоха и выдоха и т. д. Особенно сильное влияние на этот показатель оказывает гравитация.
С возрастом величина парциального давления кислорода в альвеолах практически не меняется, несмотря на значительные возрастные изменения многих показателей внешнего дыхания (уменьшение ЖЕЛ, ОЕЛ, проходимости бронхов, увеличение ФОЕ, ООЛ и т. д. ). Сохранению устойчивости показателя рО2 в альвеолах способствует возрастное увеличение частоты дыхания.
Факторы, способствующие диффузии газов в легких. Закон Фика. Диффузионная способность легких:
Диффузия газов между альвеолярным воздухом и кровью подчиняется общему закону диффузии, согласно которому ее движущей силой является разность парциальных давлений (напряжений) газа между альвеолами и кровью.
|
|
|
В притекающей к легким плазме крови газы находятся в растворенном состоянии и создают напряжение в крови, которое выражают в тех же единицах (мм рт. ст. ), что и парциальное давление.
В поступающей в капилляры малого круга крови среднее напряжение кислорода (рО2) равно 40 мм рт. ст., а его парциальное давление в альвеолярном воздухе –100 мм рт. ст. Градиент давления кислорода между альвеолярным воздухом и кровью составляет 60 мм рт. ст. В притекающей крови рСО2 – 46 мм, в альвеолах – 40 мм рт. ст. и градиент давления углекислого газа составляет 6 мм рт. ст.
Эти градиенты и являются движущей силой газообмена между альвеолярным воздухом и кровью. Следует учитывать, что указанные величины градиентов имеются лишь в начале капилляров, по мере продвижения крови по капилляру разность между парциальным давлением в альвеолярном газе и напряжением в крови уменьшается.
На скорость обмена кислорода между альвеолярным воздухом и кровью влияют как свойства среды, через которую идет диффузия, так и время (около 0, 2 с), в течение которого происходит связывание перешедшей порции кислорода с гемоглобином.
Для перехода из альвеолярного воздуха в эритроцит и связи с гемоглобином молекула кислорода должна продиффундировать через:
· 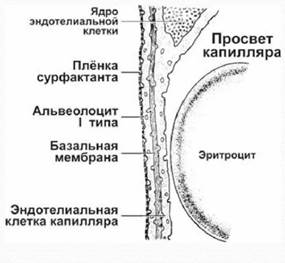 Слой сурфактанта, выстилающий альвеолу;
Слой сурфактанта, выстилающий альвеолу;
· Альвеолярный эпителий;
· Базальные мембраны и интерстициальное пространство между эпителием и эндотелием;
· Эндотелий капилляра;
· Слой плазмы крови между эндотелием и эритроцитом;
· Мембрану эритроцита;
· Слой цитоплазмы и слой гемоглобина в эритроците.
Суммарное расстояние этого диффузионного пространства составляет от 0, 5 до 2 мкм.

На практике в диагностических целях определяют показатель, называемый диффузионной способностью легких для кислорода ( ДЛО2 ).
Она равна объему кислорода, продиффундировавшему из альвеолярного воздуха в кровь через всю поверхность газообмена за 1 мин при градиенте давления кислорода 1 мм рт. ст.
 VО2 – поглощение кислорода за 1 мин;
VО2 – поглощение кислорода за 1 мин;
Po2 – разность давлений (P1 и P2)
Р1 – парциальное давление кислорода в альвеолах;
Р2 – напряжение кислорода в крови.
В норме, когда взрослый человек находится в состоянии покоя величина ДЛО2 равна 20–25 мл/мин · мм рт. ст. При физической нагрузке ДЛО2 увеличивается и может достигнуть 70 мл/мин·мм рт. ст.
|
|
|
У пожилых людей величина ДЛО2 снижается; в 60 лет она приблизительно на 1/3 меньше, чем у молодых людей.
Коэффициент проницаемости биологических сред для СО2 в 20–25 раз выше, чем у кислорода. Поэтому диффузия углекислого газа в тканях организма и в легких при меньших, чем для кислорода, градиентах его концентраций, идет быстро и углекислый газ, содержащийся в венозной крови при большем (46 мм рт. ст. ), чем в альвеолах (40 мм рт. ст. ), парциальном давлении, как правило, успевает выходить в альвеолярный воздух даже при некоторой недостаточности кровотока или вентиляции, в то время как обмен кислорода в таких условиях уменьшается.
Скорость движения крови в легочных капиллярах такая, что один эритроцит проходит через капилляр за 0, 75–1 с. Этого времени вполне достаточно для практически полного уравновешивания между парциальным давлением кислорода и углекислого газа в альвеолах и напряжением их в крови легочных капилляров, а для связывания кислорода гемоглобином эритроцита требуется лишь около 0, 2 с.
В оттекающей от легких по венам малого круга артериальной крови у здорового человека в обычных условиях напряжение кислорода составляет 85– 100 мм рт. ст., напряжение углекислого газа – 35–45 мм рт. ст.
161. Транспорт газов кровью. Соединение гемоглобина с газами.
Объем газов, находящихся в крови, принято выражать в объемных процентах (об. %). Этот показатель отражает количество газа в миллилитрах, находящееся в 100 мл крови. Кислород транспортируется кровью в состоянии физического растворения (0, 3 об. %, т. е. 0, 3 мл кислорода в 100 мл крови) и в виде химической связи с гемоглобином (15- 21 об. %). Молекулу гемоглобина, не связанную с кислородом, обозначают символом Нв, а присоединившую кислород — Нв02. Присоединение кислорода к гемоглобину называют оксигенацией, а его отдачу деоксигенациейили восстановлением. Гемоглобину принадлежит основная роль в транспорте кислорода. Одна молекула гемоглобина связывает 4 молекулы кислорода. Один грамм гемоглобина связывает и транспортирует 1, 34 мл кислорода. Зная содержание Нв в крови, легко рассчитать кислородную емкость крови.
|
|
|
Связывание О2
97% О2, транспортирующегося от легких к тканям, химически связано с гемоглобином.
Молекула гемоглобина состоит из 4 субъединиц, поэтому реакция оксигенации протекает следующим образом: Hb + 4О2 = Hb(О2)4. Молекула О2 обратимо связывается с гемом гемоглобина. Каждый грамм гемоглобина может связать 1, 34 мл О2 (число Хюфнера) (1, 39 мл, если гемоглобин химически чистый). Следовательно, при содержании гемоглобина в 1 л крови 140 г в этом объеме будет находиться (без учета физически растворённого О2) 1, 34× 140 = 190 мл О2, т. е. 19 об. %. Количество О2, которое вообще способна связать кровь, называется кислородной ёмкостью крови (в среднем 20 об. %). Реализуется полностью, если кровь контактирует с газовой смесью с высоким содержанием О2 (РО2 > 300 мм рт. ст. ). В естественных условиях гемоглобин оксигенирован только на 97%.
Насыщением (saturation) гемоглобина кислородом называется процентное отношение HbО2 к общему содержанию гемоглобина.
SО2 = [HbО2] / [Hb]+[HbО2]
Реакция оксигенации гемоглобина, как указывалось, подчиняется закону действующих масс. Это означает, что отношение между гемоглобином и HbО2 зависит от содержания физически растворенного О2 в крови, которое, в свою очередь, в соответствии с законом Генри-Дальтона, пропорционально напряжению О2. Графически эту зависимость отражает кривая диссоциации оксигемоглобина (КДО).
162. Транспорт кислорода кровью. Кислородная ёмкость крови. Насыщение крови кислородом. Факторы, способствующие оксигенации крови. Обмен О2.
Молекулы О2 диффундируют в сторону более низких величин напряжения этого газа: из капилляров (рО2 = 96 мм рт. ст. ) в межклеточную жидкость (рО2 = 20-40 мм рт. ст. ), затем в клетки.
Наименьшее напряжение О2 в клетках наблюдается в местах его потребления – митохондриях, где он используется для биологического окисления. Уровень дыхания в митохондриях остается постоянным, пока экстрамитохондриальное напряжение кислорода не упадет ниже 5 мм рт. ст. Кроме этого, О2, поступивший в клетки скелетных мышц и миокарда, связывается с миоглобином. Величина потребления О2 различна в разных тканях и связана с их активностью.
В результате отдачи О2 тканям его количество в крови постепенно уменьшается. Поэтому скорость этого процесса больше в артериальном конце капилляра, чем в венозном. Зная число Хюфнера (1, 34) и степень насыщения гемоглобина кислородом (SО2) в артериальном и венозном концах капилляра, можно рассчитать объемное содержание О2 в крови:
|
|
|
О2 = 1, 34 × [Hb] × SO2 × 10-5
где SO2 выражается в %,
[Hb ] в г/л.
В артериальной крови (SO2 = 97%) содержание химически связанного О2 около 0, 20 л на л крови; в венозной (SO2 = 75%) – 0, 15 л на л крови. Следовательно, артерио-венозная разница по О2 равна 0, 05. Это означает, что при прохождении крови через тканевые капилляры используется лишь 25% кислородной емкости. При интенсивной нагрузке артерио-венозная разница по О2 может превышать 0, 1.
Количество потребленного О2 в процентах от его содержания в артериальной крови называют коэффициентом утилизации кислорода (КУК). В норме он равен 25%, после физической нагрузки может достигать 75-85%. В локальных участках тканей, где ток крови чрезвычайно медленный или очень высока скорость метаболизма, КУК может возрастать до 100%, т. е. из крови будет поглощаться весь кислород.
Факторы, влияющие на величину КУК:
1. Количество функционирующих капилляров, их геометрия, скорость кровотока.
2. рН, температура и другие факторы, влияющие на диссоциацию HbO2.
|
|
|
