 |


Сцепленное наследование генов и кроссинговер. Работы Т. Моргана. Хромосомная теория. Примеры сцепленного наследования признаков у человека.
|
|
|
|
Сцепленное наследование — феномен скоррелированного наследования определённых состояний генов, расположенных в одной хромосоме.
Полной корреляции не бывает из-за мейотического кроссинговера, так как сцепленные гены могут разойтись по разным гаметам. Кроссинговер наблюдается в виде расцепления у потомства тех аллелей генов и, соответственно, состояний признаков, которые были сцеплены у родителей.
Наблюдения, проведённые Томасом Морганом, показали, что вероятность кроссинговера между различными парами генов разная, и появилась идея создать генные карты на основании частот кроссинговера между разными генами. Первая генная карта была построена студентом Моргана, Альфредом Стёртевантом (англ.) в 1913 году на материале Drosophila melanogaster.
Расстояние между генами, расположенными в одной хромосоме, определяется по проценту кроссинговера между ними и прямо пропорционально ему. За единицу расстояния принят 1 % кроссинговера (1 морганида или 1 сантиморганида). Чем дальше гены находятся друг от друга в хромосоме, тем чаще между ними будет происходить кроссинговер. Максимальное расстояние между генами, расположенными в одной хромосоме, может быть равно 49 сантиморганидам.
Сцепленное наследование генов
Мендель изучил наследование только семи пар признаков у душистого горошка. Его законы подтвердились на самых разных видах организмов, т. е. было признано, что эти законы носят всеобщий характер. Однако позже было замечено, что у душистого горошка два признака — форма пыльцы и окраска цветков — не дают независимого распределения в потомстве. Потомки оставались похожими на родителей. Постепенно таких исключений из третьего закона Менделя накапливалось все больше. Стало ясно, что принцип независимого распределения в потомстве и свободного комбинирования распространяется не на все гены. Действительно, у любого организма признаков очень много, а число хромосом невелико.
|
|
|
В каждой хромосоме должно локализоваться много генов. Каковы же закономерности наследования генов, локализованных & одной хромосоме? Вопрос этот был изучен выдающимся американским генетиком Т. Морганом.
Предположим, что два гена — А и В находятся в одной хромосоме и организм, взятый для скрещивания, гетерозиготен по этим генам:
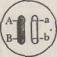
В анафазе I мейотического деления гомологичные хромосомы расходятся к разным полюсам и образуются два типа гамет
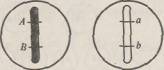
вместо четырех, как должно быть при дигибридном скрещивании в соответствии с третьим законом Менделя.
При скрещивании с организмом, рецессивным по обоим генам aabb, получается расщепление 1:1

вместо ожидаемого при дигибридном анализирующем скрещивании 1:1:1:1. Такое отклонение от независимого распределения означает, что гены, локализованные в одной хромосоме, наследуются совместно.
Явление совместного наследования генов, локализованных в одной хромосоме, называется сцепленным наследованием, а локализация генов в одной хромосоме — сцеплением генов. Сцепленное наследование генов, локализованных в одной хромосоме, установил Морган.
Таким образом, третий закон Менделя применим лишь к наследованию аллельных пар, находящихся в негомологичных хромосомах.
Все гены, входящие в состав одной хромосомы, передаются по наследству совместно и составляют группу сцепления. Поскольку в гомологичных хромосомах находятся одинаковые гены, группу сцепления составляют две гомологичные хромосомы. Число групп сцепления у данного вида организмов соответствует числу хромосом в гаплоидном наборе. Так, у человека 46 хромосом в диплоидном наборе — 23 группы сцепления, у дрозофилы 8 хромосом — 4 группы сцепления, у гороха 14 хромосом — 7 групп сцепления. Однако при анализе наследования сцепленных генов было обнаружено, что в определенном проценте случаев сцепление может нарушаться.
|
|
|
Вспомним, что в профазе I мейотического деления гомологичные хромосомы конъюгируют. В этот момент может произойти обмен участками гомологичных хромосом:

Предположим, что в одной из гомологичных хромосом локализуются пять известных нам доминантных генов, а в другой — пять их рецессивных аллелей. Если проследить распределение в потомстве двух генов — А и В, то в результате расхождения гомологичных хромосом в анафазе I мейотического деления дигетерозиготный организм в случае сцепления генов А и В должен давать два типа гамет: АВ и ab. Но если в результате кроссин-говера в некоторых клетках происходит обмен участками хромосом между генами А и В, то появляются гаметы АЬ и аВ, и в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Отличие заключается в том, что числовое отношение фенотипов не соответствует отношению 1:1:1:1, установленному для дигибридного анализирующего скрещивания.
Таким образом, сцепление генов может быть полным и неполным. Причина нарушения сцепления — кроссин-говер, т. е. перекрест хромосом в профазе I мейотического деления. Чем дальше друг от друга расположены гены в хромосоме, тем выше вероятность перекреста между ними и тем больше процент гамет с перекомбинированными генами. В генетике принято определять расстояние между генами в процентах гамет, при образовании которых в результате кроссинговера произошла перекомбинация генов в гомологичных хромосомах. Кроссинговер — важный источник комбинативной генетической изменчивости.
Работы Моргана заложили основы хромосомной теории наследственности, они показали, что ограничения в свободной комбинаторике некоторых генов обусловлены расположением этих генов в одной хромосоме и их физическим сцеплением.
Морганом было установлено, что сцепление генов, расположенных в одной хромосоме, не является абсолютным. Во время мейоза хромосомы одной пары могут обмениваться гомологичными участками между собой с помощью процесса, который называется кроссинговером. Чем дальше друг от друга расположены гены в хромосоме, тем чаще они разделяются кроссинговером. На основе этого феномена была предложена мера силы сцепления генов — процент кроссинговера — и построены первые генетические карты хромосом для разных видов дрозофилы. У дрозофилы гигантские хромосомы слюнных желез представляли собой не только идеальный объект для цитологического изучения. Морган и его сотрудники использовали хромосомные мутации как цитологические маркеры расположения генов. Собственно такое совмещение цитологического и генетического изучения хромосом и создало особый раздел генетики, который называется цитогенетикой. Чтобы стало ясным, насколько важным для генетики всех высших организмов является установление точного расположения генов в хромосомах, можно указать, что в Проекте «Геном человека» создание точных генетических карт было и даже остается одним из основных направлений исследований. Впервые предположение о том, что хромосомы являются носителями наследственной информации в клетке, было высказано еще в 1902 г. Т.Бовери, В.Сэттоном и К.Коррен-сом, но оно основывалось на цитологических доказательствах поведения хромосом во время деления клеток.
|
|
|
|
|
|
