 |


А – нормальная хромосома, Б – делетированная хромосома
|
|
|
|


А Б
Рис. 14.3.5. Синдром "кошачьего крика".
А – дети с синдромом Лежена в возрасте 8 месяцев, 2, 4 и 8 лет соответственно;
Б - хромосомный набор больного с синдромом Лежена: групповая (от А до G) и индивидуальная идентификация хромосом (стрелкой указан дефект короткого плеча хромосомы 5-й пары, вторая хромосома не изменена).
- дупликации (от лат. duplcatio - удвоение) - удвоение (или умножение) какого-либо участка хромосомы(например, трисомияпо одному из коротких плеч 9-й хромосомы обусловливает множественные пороки, включая микроцефалию, задержку физического, психического и интеллектуального развития) (рис. 14.3.6);

Рис. 14.3.6. Схематическое изображение дупликации.
А – нормальная хромосома, Б – дуплицированная хромосома.
- инверсии (от лат. inversio - перевертывание). В результате двух точек разрывов хромосомы образовавшийся фрагмент встраивается на прежнее место после поворота на 180°. В результате нарушается только порядок расположения генов (рис. 14.3.7);

Рис. 14.3.7. Схематическое изображение инверсии.
А – нормальная хромосома, Б – инвертированная хромосома.
Межхромосомные мутации, илимутации перестройки - обмен фрагментами между негомологичными хромосомами. Такие мутации получили название транслокации (от лат. trans - за, через + locus - место) (рис. 14.3.8).
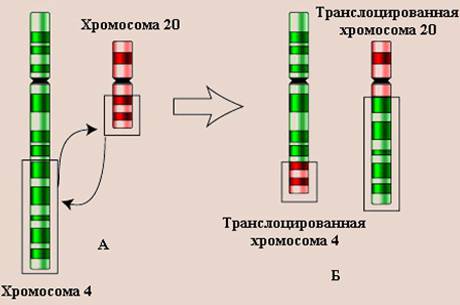
Рис. 14.3.8. Схематическое изображение транслокации хромосом.
А – нормальные хромосомы, Б – транслоцированные хромосомы.
Это:
- реципрокная транслокация, когда две хромосомы обмениваются своими фрагментами;
- нереципрокная транслокация, когда фрагмент одной хромосомы транспортируется на другую;
- "центрическое" слияние (робертсоновская транслокация) – соединение двух акроцентрических хромосом в районе их центромер с потерей коротких плеч.
|
|
|
При поперечном разрыве хроматид через центромеры "сестринские" хроматиды становятся "зеркальными" плечами двух разных хромосом, содержащих одинаковые наборы генов. Такие хромосомы называют изохромосомами.
Как внутрихромосомные (делеции, инверсии и дупликации), так и межхромосомные (транслокации) аберрации и изохромосомы связаны с физическими изменениями структуры хромосом, в том числе с механическими разломами.
Геномные
К геномным мутациям относятся анеуплоидии и изменение плоидности структурно неизмененных хромосом. Выявляются цитогенетическими методами.
Геномные мутации в животном и растительном мире многообразны, но у человека обнаружены только 3 типа геномных мутаций: тетраплоидия, триплоидия и анеуплоидия. При этом из всех вариантов анеуплоидий встречаются только трисомии по аутосомам, полисомии по половым хромосомам (три-, тетра- и пентасомии), а из моносомий встречаются только моносомия-Х.
Анеуплоидия - изменение (уменьшение - моносомия, увеличение - трисомия) числа хромосом в диплоидном наборе, некратное гаплоидному (2n + 1, 2n - 1 и т.д.).
Полиплоидия - увеличение числа наборов хромосом, кратное гаплоидному (Зn, 4n, 5n и т.д.).
У человека полиплоидия, а также большинство анеуплоидий являются летальными мутациями.
К наиболее частым геномным мутациям относятся:
трисомия - наличие трех гомологичных хромосом в кариотипе (например, по 21-й паре, при синдроме Дауна, по 18-й паре при синдроме Эдвардса, по 13-й паре присиндроме Патау; по половым хромосомам: XXX, XXY, XYY) (рис. 14.3.8,);
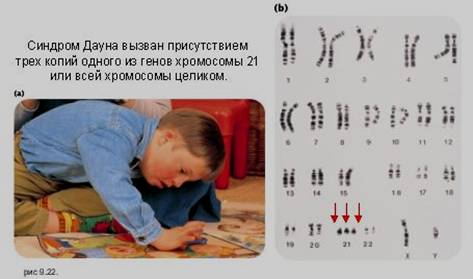
Рис. 14.3.8. Подавляющая часть случаев синдрома Дауна вызвана нерасхождением пары хромосом 21 в мейозе у матери;
а – ребенок, больной синдромом Дауна, б – стрелки указывают на три копии генов хромосомы 21 (47, ХХ + 21).
моносомия - наличие только одной из двух гомологичных хромосом. При моносомии по любой из аутосом нормальное развитие эмбриона невозможно. Единственная моносомия у человека, совместимая с жизнью, - моносомия по X-хромосоме - приводит к синдрому Шерешевского-Тернера (45, Х0) (рис. 14.3.9).
|
|
|

Рис. 14.3.9. Девочка, больная синдромом Тернера (слева).
Складки кожи в области шеи — характерный признак болезни.
Справа хромосомы человека с синдромом Тернера (45, Х0)
Причиной, приводящей к анеуплоидии, является нерасхождение хромосом во время клеточного деления при образовании половых клеток или утрата хромосом в результате анафазного отставания, когда во время движения к полюсу одна из гомологичных хромосом может отстать от всех других негомологичных хромосом. Термин "нерасхождение" означает отсутствие разделения хромосом или хроматид в мейозе или митозе. Утрата хромосом может приводить к мозаицизму, при котором имеется одна эуплоидная (нopмальная) клеточная линия, а другая - моносомная.
Нерасхождениехромосом наиболее часто наблюдается во время мейоза (рис. 14.3.10). Хромосомы, которые в норме должны делиться во время мейоза, остаются соединенными вместе и в анафазе отходят к одному полюсу клетки. Таким образом, возникают две гаметы, одна из которых имеет добавочную хромосому, а другая не имеет этой хромосомы. При оплодотворении гаметы с нормальным набором хромосом гаметой с лишней хромосомой возникает трисомия (т.е. в клетке присутствуют три гомологичные хромосомы), при оплодотворении гаметой без одной хромосомы возникает зигота с моносомией. Если моносомная зигота образуется по какой-либо аутосомной (не половой) хромосоме, то развитие организма прекращается на самых ранних стадиях развития.
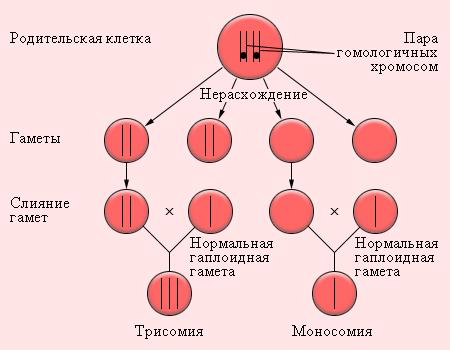
Рис. 14.3.10. Схема нерасхождения хромосом в гаметогенезе и результаты оплодотворения
Летальный или дизморфогенетический эффект хромосомных аномалий обнаруживается на всех стадиях внутреутробного онтогенеза (имплантация, эмбриогенез, органогенез, рост и развитие плода). Суммарный вклад хромосомных аномалий во внутриутробную гибель (после имплантации) у человека составляет 45 %. При этом, чем раньше прерывается беременность, тем вероятнее, что это обусловлено аномалиями развития эмбриона, вызванными хромосомным дисбалансом. У 2-4-недельных абортусов (эмбрион и его оболочки) хромосомные аномалии обнаруживают в 60 - 70 % случаев. В 1 триместре беременности хромосомные аномалии встречаются у 50 % абортусов. У плодов-выкидышей 2 триместра такие аномалии находят в 25 - 30 % случаев, а у плодов, погибших после 20 нед. беременности, в 7 % случаев.
|
|
|
Наиболее тяжелые формы по дисбалансу хромосомного набора встречаются у ранних абортусов. Это полиплоидии (25 %), полные трисомии по аутосомам (50 %). Трисомии по некоторым аутосомам (1; 5; 6; 11; 19) встречаются крайне редко даже у элиминированных эмбрионов и плодов, что свидетельствует о большой морфогенетической значимости этих аутосом. Данные аномалии прерывают развитие в доимплантационном периоде или нарушают гаметогенез.
Высокая морфогенетическая значимость аутосом еще более отчетливо выражена при полных аутосомных моносомиях. Последние редко обнаруживаются даже в материале ранних спонтанных абортов из-за раннего летального эффекта такого дисбаланса.
Типы мутаций
Мутации являются начальным звеном патогенеза наследственных болезней.
По виду клеток, в которых произошли изменения, мутации можно разделить на:
гаметические (от греч. gamete - супруга), или генеративные, мутации в половых клетках. Они наследуются и, как правило, обнаруживаются во всех клетках потомков, ставших их носителями;
соматические - мутации в неполовых клетках организма. Проявляются у того индивида, у которого они возникают. Они передаются только дочерним клеткам при делении и не наследуются следующим поколением индивида. Если соматическая мутация возникает на ранних стадиях дробления зиготы (но не первого деления), возникают клеточные линии с различными генотипами. Чем раньше в онтогенезе происходит соматическая мутация, тем больше клеток и соответственно тканей несет данную мутацию. Подобные организмы получили название мозаичных. По влиянию на организм можно выделить:
1) летальные мутации - это мутации, которые приводят к внутриутробной гибели или к смерти в младенческом возрасте. Например, такая геномная мутация, как моносомия по аутосомам, учеловека несовместима с нормальным развитием эмбриона;
|
|
|
2) полулетальные мутации - мутации, значительно снижающие жизнеспособность организма, приводя к ранней смерти. Продолжительность жизни носителей полулетальных мутаций может значительно варьировать, однако в любом случае они погибают до достижения половой зрелости (например, при пигментной ксеродерме);
3) нейтральные мутации - мутации, не влияющие существенным образом на процессы жизнедеятельности;
4) благоприятные мутации - мутации, обеспечивающие организму новые полезные свойства.
В соответствии с уровнем организации наследственных структур различают генные, хромосомные и геномные мутации (рис. 14.3.1).
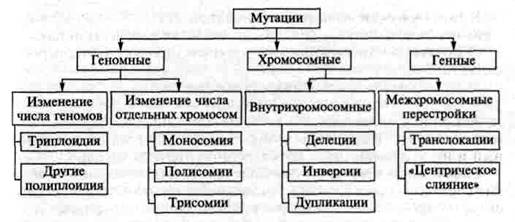
Рис. 14.3.1. Схема типов мутаций
Геномные и хромосомные мутации являются причинами возникновения хромосомных болезней.
Номенклатура мутаций
В 1992 г. американскими учеными Артуром Боде и Лап-Чи Тсуи была предложена универсальная стандартная система для обозначения разных мутаций. Она рассчитана как на запись аминокислотных замен в белках, так и на нуклеотидные замены и перестановки в ДНК. В первом случае каждой аминокислоте соответствует однобуквенный символ (табл. № 14.4.1), слева записывается нормальный вариант аминокислоты, справа - мутантный, а расположенный в центре номер соответствует месту замены в цепочке первичного продукта трансляции.
Таблица № 14.4.1
Обозначение аминокислот
| Трехбуквенное обозначение | Аминокислота | Однобуквенное обозначение | Трехбуквенное обозначение | Аминокислота | Однобуквенное обозначение |
| Ala | Аланин | (A) | Leu | Лейцин | (L) |
| Arg | Аргинин | (R) | Lys | Лизин | (K) |
| Asn | Аспарагин | (N) | Met | Метионин | (M) |
| Asp | Аспарагин.к-та | (D) | Phe | Фенилаланин | (F) |
| Cys | Цистеин | (C) | Pro | Пролин | (P) |
| Gln | Глутамин | (Q) | Ser | Серин | (S) |
| Glu | Глутамин.к-та | (E) | Thr | Треонин | (T) |
| Gly | Глицин | (G) | Trp | Триптофан | (W) |
| His | Гистидин | (H) | Tyr | Тирозин | (Y) |
| Ile | Изолейцин | (I) | Val | Валин | (V) |
Например, запись N44G означает замену аспарагина на глицин в 44-м положении полипептидной цепи, а А6559 - аланина на глутамин в положении 655 белкового продукта. Так записываются различные варианты аминокислотных замен при миссенс-мутациях. Буквой Х обозначается место остановки синтеза полипептидной цепи при нонсенс-мутациях. Например, G39X означает замену глицина на стоп-сигнал в 39-м кодоне, a W1282X - триптофан-триплета на стоп-кодон в положении 1282. Отсутствие одной или нескольких аминокислот обозначают значком дельта. Так, наиболее частая мутация, приводящая к муковисцидозу, дельтаF508 - означает отсутствие фенилаланина в 508-м положении трансмембранного регуляторного белка муковисцидоза.
|
|
|
Полиморфизмы, связанные с равноценной по функциональной значимости заменой аминокислот, записывают через косую черту. Например, M/V470 - метионин или валин в положении 470.
Принципиальная схема записи и нумерации нуклеотидов приведена на рис. 14.4.1.
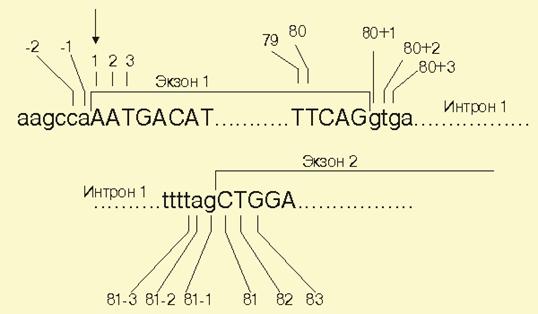
Рис. 14.4.1. Принцип нумерации нуклеотидов в гене.
Отсчет нуклеотидов в молекуле ДНК начинается с первого смыслового кодона, так что нуклеотид под номером +1 соответствует первому нуклеотиду в молекуле кДНК. Вверх по течению (или справа налево от 3' к 5'-концу) от первого кодона нуклеотиды записывают со знаком "-", вниз по течению (от 5' - к 3') - со знаком "+". Для многих генов отсутствие точных данных о положении инициирующего сайта и наличие нескольких мест инициации транскрипции существенно затрудняют нумерацию нуклеотидов.
Нуклеотиды экзонов обозначают заглавными буквами, интронов - прописными.
В табл. № 14.4.2 даны примеры обозначения различных мутаций с использованием как аминокислотной, так и нуклеотидной нумерации.
Таблица № 14.4.2
Примеры обозначения мутаций в гене муковисцидоза
| Название | Изменения в нуклеотидах | Позиция | Изменения в аминокислотах | Позиция | Экзон |
| Миссенс | |||||
| D44G | A-G | Asp-Gly | |||
| A455E | C-A | Ala-Glu | |||
| S549(A-C) | A-C | Ser-Arg | |||
| S549(T-G) | T-G | Ser-Arg | |||
| Нонсенс | |||||
| Q39X | C-T | Gln-Stop | |||
| W1282X | G-A | Trp-Stop | |||
| Вставка, делеция, сдвиг рамки считывания | |||||
| delF508 | Делеция 3 п.о. | 1652-1655 | Делеция F | ||
| 241delAT | Делеция AT | Сдвиг рамки | |||
| 852del22 | Делеция 22 п.о. | Сдвиг рамки | 6a | ||
| 1154insTC | Вставка TC | Сдвиг рамки | |||
| Сплайсинг | |||||
| 621+1G-T | G-T | 621+1 | 5`-сигнал | Интрон 4 | |
| 622-2A-C | A-C | 622-2 | 3`-сигнал | Интрон 4 | |
| 3849+10kbC-T | C-T | Ошибочный сплайсинг | Интрон 19 | ||
| Полиморфизмы | |||||
| M/V470 | AG | M или G | |||
| 1716G/A | GA | Без изменений | |||
| 125G/C | GC | 5`-нетранслируемаяя область | - | - |
Нуклеотидная система записи особенно важна для обозначения делеций, инсерций, сплайсинговых мутаций и полиморфизмов, не связанных с заменами аминокислот или происходящих в нетранслируемых частях гена. В случае делеции или инсерций одного или двух нуклеотидов приводится их буквенное обозначение. Например, 441delA, 485insTA. При делеции или инсерции трех и более нуклеотидов указывается только их число. Так, 852del22 означает делению 22 нуклеотидов, начиная с 852-го нуклеотида, a 3320ins7 обозначает вставку 7 пар оснований после нуклеотида 3320. В случае больших вставок или делеций их размеры указываются в килобазах, например, 2115 ins 13kb, или обозначаются соответствующие инсертированные/делетированные структурные элементы генома. Так, 2115insAlu означает инсерцию Alu-повтора после нуклеотида 2115.
При обозначении сплайсинговых мутаций записывают номер крайнего нуклеотида в ближайшем к мутации экзоне, число нуклеотидов (со знаком "+" в случае 3'-конца экзона и со знаком "-" в случае 5'-конца) и характер нуклеотидной замены. Например, запись 711+5G-T обозначает замену G на Т в 5-м основании интрона, следующего за экзоном, заканчивающимся 711-м нуклеотидом.
|
|
|
