 |


Голондрическое наследование
|
|
|
|
Так как гетерогаметный пол гемизиготен по сцепленным с полом генам, то эти гены всегда проявляются в их фенотипе, даже если они рецессивны. Большинство генов, имеющихся в Х-хромосоме, в Y-хромосоме отсутствует, однако определенную генетическую информацию она все-таки несет. Различают два типа такой информации: во-первых, содержащуюся в генах, присутствующих только в Y- хромосоме, и, во-вторых, в генах, присутствующих как в Y-, так и в Х-хромосоме (геморрагический диатез).
Y-хромосома передается от отца всем его сыновьям, и только им. Следовательно, для генов, содержащихся только в Y-хромосоме, характерно голондрическое наследование, т.е они передаются от отца к сыну и проявляются у мужского пола (рис. 12.3.).
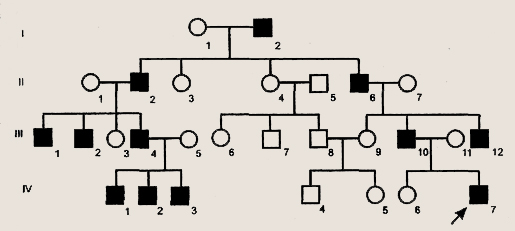
Рисунок 12.3. Пример родословной с голондрическим типом наследования
Долгое время полагали, что Y -хромосома человека, как и некоторых других живых организмов, например дрозофилы, является "пустой", или "генетически инертной", т.е. считалось, что в Y -хромосоме отсутствуют какие-либо гены, ответственные за развитие тех или иных признаков.
В настоящее время установлено, что по длине Y -хромосомы у человека локализовано по меньшей мере 10 генов, среди которых, например, ген гипертрихоза (оволосение мочки уха) и ген синдактилии (перепончатости, т.е. сращения пальцев рук, например, указательного и среднего). Ген, ответственный за развитие такого признака, как гипертрихоз (оволосение края мочки уха), – один из немногих рецессивных генов, локализованных в Y-хромосоме. Если мужчина с гипертрихозом женится на женщине, у которой, естественно, гипертрихоза нет, то каков реальный шанс появления в этой семье детей с гипертрихозом, как мальчиков, так и девочек?
|
|
|

Все просто: 100% девочек, рожденных от этого мужчины, здоровы; все мальчики – с гипертрихозом.
Наследование признаков, контролируемых полом
Кроме признаков, сцепленных с полом, имеются такие признаки, которые контролируются и ограничиваются полом. Ряд признаков организма, расположенных в аутосомах, имеют разную степень проявления у мужских и женских особей. Признаки, выражение или проявление которых различно у представителей разных полов, или проявляющиеся только у одного пола, относятся к ограниченным полом. Степень проявления аутосомных генов контролируется половыми гормонами, которые являются стимуляторами некоторых признаков. Например, у мужчин ген облысения проявляется как доминантный в гетерозиготном состоянии, а у женщин – это рецессивный ген и проявляется только в гомозиготном состоянии. Появление лысины - аутосомно-доминантный признак, но проявляется практически только у мужчин при наследовании, контролируемом полом, у женщин подавляются гены, детерминирующие рост бороды. Другим примером признака, ограниченного полом, является молочность коров и жирность молока. Эти признаки имеются и у быков, но проявляются только у коров
Геномный импринтинг
Эпигенетика занимается изучением механизмов наследственности, которые модифицируют работу генов без изменения их структуры. Причем эти модификации сохраняются при митотическом делении клеток. Наиболее часто эпигенетические механизмы обеспечивают долговременную репрессию значительной части генома в специализированных клетках многоклеточного организма. Эпигенетические механизмы обеспечивают и импринтинг – инактивацию ряда генов в одной из родительских хромосом. Наиболее изученным механизмом является метилирование ДНК.
При импринтинге специфический характер дифференциальной активности генов определяется полом организма, от которого эти гены унаследованы. У некоторых насекомых, например, грибных комариков (Sciaridae), весь набор отцовских хромосом элиминируется во время сперматогенеза. У этих организмов отцовские хромосомы маркируются в цитоплазме клеток зародышевой линии, удаляются при созревании гамет и не передают свои гены следующему поколению. У млекопитающих и высших цветковых растений отцовские и материнские гены оказывают разный эффект на развитие эмбриона, но в одинаковой степени представлены в гаметах, образующихся в результате мейоза. В этом случае вклады отцовского и материнского геномов в развитие организмов не эквивалентны и происходит видимое искажение менделевских правил наследования некоторых признаков. Иными словами, характер экспрессии отцовского и материнского аллелей одного и того же гена в организме-потомке может быть различным, так как зависит от их происхождения. Молекулярные механизмы этого явления в настоящее время до конца непонятны. Предполагается, что в развитие феномена импринтинга вносят вклад зависимые от пола модификации ДНК определенных генов, в частности метилирование их регуляторных участков.
|
|
|
Импринтинг распространяется на узкую группу (более 20) генов млекопитающих, отличающихся от большинства генов тем, что их аллели функционально неравноценны. Обычно копии того или иного гена либо обе не работают, либо равно активны. В случае импринтированных генов активен только один аллель (отцовский или материнский), причем статус строго детерминирован именно происхождением (от какой из родительских клеток).
Суть импринтинга в том, что гены, передаваемые потомству, несут специфический "отпечаток" пола родителя, т. е. отцовские и материнские гены маркированы по-разному; причем эти "отпечатки" временные и могут быть "стерты". Вследствие геномного импринтинга потомки, получившие маркированные гены от матери, отличаются от тех, которые унаследовали такие гены от отца. Другими словами, в некоторых случаях важно, от которого из родителей унаследован ген. Геномный импринтинг может затрагивать любой ген. В основе исключений, связанных со сцеплением с полом или с наследованием органелл по материнской линии, лежат различия в генетическом вкладе родителей в генотип потомков. В случае геномного импринтинга, напротив, оба родителя могут передавать потомкам совершенно идентичные гены, но при разном - материнском либо отцовском - импринтинге действие этих генов будет неодинаковым (рис. 12.4).
|
|
|

Рисунок 12.4. Этапы метилирования и деметилирования генов
Геномный импринтинг состоит в том, что хромосомы половых клеток (сперматозоидов или яйцеклеток) индивида приобретают "отпечаток" его пола. Потомство получает один набор хромосом с отцовской маркировкой некоторых генов, а другой - с материнской. При образовании у потомка половых клеток прежний "отпечаток" стирается и эти гены маркируются в соответствии с полом данной особи (рис. 12.4.). Был разработан микрохирургический метод переноса клеточных ядер мышиных эмбрионов, который позволил физически заменять генетическую информацию одной особи на аналогичную информацию от другой особи. Этот метод основан на том, что после оплодотворения яйцеклетки сперматозоидом зигота некоторое время не делится и ядра яйцеклетки и сперматозоида существуют в ее цитоплазме автономно друг от друга. С помощью тончайших стеклянных капилляров из оплодотворенной яйцеклетки избирательно удаляли ядро либо яйцеклетки, либо сперматозоида (либо оба) (рис. 12.5.). Затем вместо удаленного ядра в клетку вводили ядро из другой оплодотворенной яйцеклетки. Было установлено, что если заменяется ядро яйцеклетки на ядро другой яйцеклетки или же ядро сперматозоида - на ядро другого сперматозоида, то эмбрионы развиваются полностью и выросшие из них особи неотличимы от нормальных мышей той же линии.
Если развитие эмбриона зависит только от набора его генов, то теоретически не имеет значения, что особь получает все гены от одного родителя, а не от двух, т. е. гиногенетические, андрогенетические и нормальные мыши должны развиваться одинаково. В действительности эти три группы животных развивались совсем не одинаково. Эмбрионы с двумя наборами хромосом от одной и той же родительской особи - самца или самки - не развиваются до конца. Обычно их развитие останавливается на ранней стадии, когда тело эмбриона состоит всего из нескольких десятков клеток. Если же развитие все-таки продолжается, что иногда случается, то у гиногенетических и андрогенетических особей имеют место поразительно различные аномалии.
|
|
|
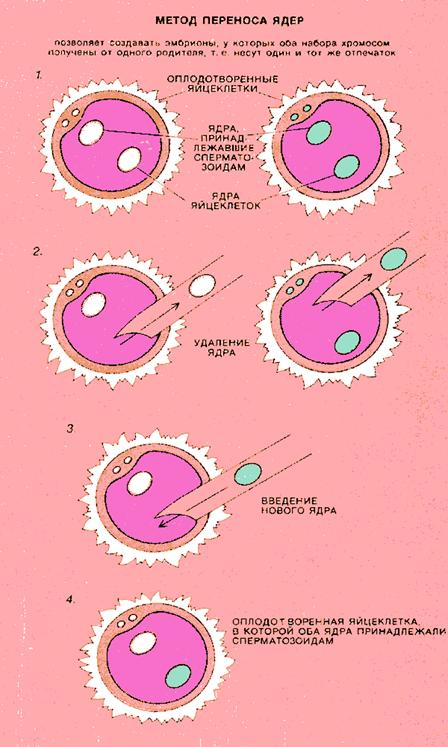
Рисунок 12.5. Метод переноса клеточных ядер
У гиногенетических эмбрионов, достигших наибольшего развития, в теле эмбриона наблюдались незначительные аномалии, однако их плаценты и желточные мешки (эти структуры необходимы для питания эмбриона) оказались сильно недоразвитыми. А у наиболее развитых андрогенетических эмбрионов, наоборот, желточные мешки и плаценты были почти нормальными, а тела - мелкими и слабо развитыми. Поскольку у гиногенетических, андрогенетических и нормальных эмбрионов последовательности нуклеотидов хромосомной ДНК не различались, было сделано заключение, что гены были каким-то образом модифицированы и несли "отпечаток" пола той особи, от которой они унаследованы. По-видимому, в результате этого геномного импринтинга избирательно инактивируются определенные гены, которые в норме функционируют в ранний период эмбрионального развития. Среди генов, унаследованных от отца, не проявляли активности некоторые гены, управляющие эмбриональным развитием; гены, важные для образования желточного мешка и плаценты, оказывались неактивными, если были получены от матери.
Явление геномного импринтинга связано с некоторыми заболеваниями человека. У многих больных с синдромов Прадера-Вилли (при этом заболевании характерны умственная отсталость, крайнее ожирение, низкий рост и непропорционально малый размер рук и ног) обе хромосомы 15 унаследованы от матери. Р. Николлс, Дж. Кнолл (Гарвардский университет) и Ч. Уильямс из Флоридского университета обнаружили связанную с геномным импринтингом закономерность у больных с синдромом Ангельмана, при котором наблюдается неадекватная смешливость, резкие судорожные движения и другие симптомы двигательной и умственной отсталости. У таких больных нередко имеют место частичные делеции унаследованной от матери хромосомы 15, в результате чего полностью функциональна только отцовская хромосома 15. Эти два заболевания, хотя и столь разные по клинической картине, могут быть связаны с различиями в импринтинге одних и тех же генов одной и той же хромосомы. Однако в отличие от ненормально крупных и мелких мышей в экспериментах Кэттанака, синдромы Прадера-Вилли и Ангельмана не удается представить просто как две стороны одной медали, т. е. объяснить избытком либо недостатком продуктов одних и тех же генов.
|
|
|
Интересный пример признака, который модифицируется зависимым от пола геномным импринтингом, представляет болезнь Гентингтона. Это смертельное неврологическое расстройство наследуется как доминантный признак, т. е. оно проявляется у каждого человека, унаследовавшего ген, обусловливающий заболевание, хотя бы от одного из родителей. Обычно болезнь боражает людей немолодых; в среднем диагноз ставится в возрасте 38 лет. Однако приблизительно в 10% случаев болезнь развивается еще до достижения зрелости и бывает, что она начинается у ребенка уже в два с половиной года. Ранее было установлено, что примерно 90% больных детей получают ген, обусловливающий заболевание, от отца. Ясно, что в этих случаях и дети, и их отцы несут один и тот же ген, ответственный за развитие болезни; однако его модификация в результате отцовского импринтинга приводит к тому, что заболевание проявляется в значительно более раннем возрасте.
Для объяснения генетики болезни Хантингтона выдвигались разнообразные гипотезы, однако лучше всего имеющиеся данные согласуются с моделью, предложенной Ч.Лэйрдом из Вашингтонского университета. Нововведение Лэйрда заключается в том, что он провел аналогию с особым классом признаков дрозофилы, характеризующихся изменчивой экспрессией, или мозаичным эффектом положения. Необычность этих признаков в том, что в отличие от большинства других признаков, обусловленных мутациями, они проявляются не во всех клетках ткани. Ткани в таких случаях оказываются как бы мозаичными: часть клеток в них имеет нормальный, а часть мутантный фенотип. Доля клеток, в которых экспрессируется мутация, определяется действием генов-модификаторов, в зависимости от которого баланс мозаицизма может сдвигаться от почти полностью нормальных до почти полностью мутантных тканей. Лэйрд предположил, что если различия в возрасте начала болезни Гентингтона отражают мозаицизм в экспрессии гена, обусловливающего заболевание, то этот мозаицизм, вероятно, зависит от генов-модификаторов. Если ген-модификатор действует так, что баланс мозаицизма сдвигается в сторону мутантных клеток и возникает почти полностью мутантная ткань, то заболевание развивается рано; при противоположном действии гена оно проявляется в позднем возрасте.
Тот факт, что при наследовании гена, обусловливающего заболевание, от отца, в большинстве случаев болезнь начинается в раннем возрасте, Лэйрд объяснил расположением гена-модификатора в Х-хромосоме. Поскольку мужчины имеют по одной X-хромосоме, любая аномалия гена-модификатора будет проявляться, а не компенсироваться, как у женщин, имеющих по две Х-хромосомы. Поэтому для мужчины больше вероятность того, что у его потомков баланс мозаицизма окажется сдвинутым в сторону большей доли мутантных клеток в тканях и болезнь разовьется в раннем возрасте. В 10% случаев раннего развития болезни Гентингтона ребенок получает соответствующий ген от матери, а не от отца; этого и следует ожидать, так как хотя и редко, но должны встречаться женщины, у которых ген-модификатор аномален в обеих Х-хромосомах.
|
|
|
