 |


Класс полихета
|
|
|
|
Тело Ро1усНае1а одето однослойным кожным эпителием, который выделяет тонкую кутикулу. Эпителий местами может быть ресничным, особенно у некоторых наиболее примитивных форм. Так, у Рго(ос1гИи5 ресничками покрыта продольная полоса на брюшной стороне тела, а кроме того, каждый сегмент опоясан двойным венчиком ресничек: на головном участке один из венчиков лежит впереди рта, как у личинки кольчецов — трохофоры (с. 263). Часто эпителий содержит железистые клетки. У сидячих полихет выделения кожных желез могут затвердевать, образуя вокруг тела прозрачную, как- бы роговую, или хитиноидную, трубочку. Часто червь прикрепляет к органической основе трубки посторонние частицы, например, песчинки, обломки раковин моллюсков, увеличивающие ее прочность (см. рис. 211, Е). Есть, наконец, формы, у которых органическая основа трубки пропитывается углекислой известью
Класс олигохета
Кожный эпителий, образующий на поверхности тонкую эластичную кутикулу, богат слизистыми железистыми клетками. Особенно многочисленны слизистые и белковые одноклеточные железы в области пояска, который в период размножения червей ясно виден
Класс гирудинеа
Тело одето снаружи довольно плотной кутикулой; лежащий под ней эпителий очень богат слизистыми железистыми клетками, а у основания эпителиальных клеток расеяны многочисленные пигментные клетки с зернистостью различного цвета. Присутствие этих клеток обусловливает окраску пиявок.
75. Мезохил, мезоглея и паренхима: сравнительный анализ.
Соединительная ткань представляет собой особый компартмент в теле животного. Компартментом соединительной ткани следует назвать следующие структурные элементы: у трихоплакса и губок-мезохил, у коралловых полипов, медуз и гребнеков — мезоглея, у высших животных — соединительная ткань или паренхима. (Мезенхима — подвижные амебоидные клетки возникающие в эпителии и мигрирующих была цель зародыша).
|
|
|
Трихоплакс Мезохил фибриллярные клетки с отркостками позволяют метабол движение. Фагоцитируют частицы становясь амебоидными
Мехохил — бесструктурный межклеточный матрикс (продукт выделения клеток) между пинако- и хоанодермой губок, состоящая из коллагеновых волокон и отдельных клеточных элементов:
Колленциты - звездчатые опорные клетки, их отростки образуют сеть;
Склероциты - скелетные клетки, образуют минер. иглы - спикулы;
Амебоциты - подвижные амебоидные клетки, фагоцитируют, обеспеч. транспорт питательных в-в и внутрикл пищеварение;
Спонгиобласты - образуют органический скелет;
Археоциты - тотипотентные клетки, много лизосом, пищеварение. Могут давать начало другим клеткам, в том числе половым;
Сперматоциты, ооциты - предшественники половых клеток;
Миоциты
Функция мезохила - опорная и транспортная
Мезоглея - студенистый внеклеточный матрикс, между эпидермисом и гастродермисом у кишечнополостных (Cnidaria) и гребневиков (Ctenophora).
Мезоглея напоминает мезохил губок, содержит коллаген, часто амебоидные клетки. Подобно мезохилу губок, часто содержит клетки, мигрировавшие из эпителиальных пластов.
Функции мезоглеи опорная (гидроскелет), трофическая (обеспечивает поступл. пит. веществ к мышцам нервам и половым клеткам). Часто представляет собой лишь тонкую прослойку между эпителиями и выполняет функции базальной пластинки. Искл: у коралловых полипов, мезоглея которых может быть обширной и содержать мышечные волокна, а также клетки, выделающие минеральный или органический скелет.
Паренхима - внеклеточная внутренняя среда. Мезодермальное происхождение
|
|
|
Пространство между органами заполнено паренхимой (мезенхимой) — тканью мезодермального происхождения. Она мало дифференцирована, практически однородна и состоит из отростчатых клеток неправильной формы. Клетки переплетаются отростками, и таким образом формируется сетчатая структура. Пространства между клетками заполнены жидкостью.
Функции паренхимы:
- запасные питательные вещества (гликоген, жиры);
- продукты метаболизма, выводящиеся затем по каналам протонефридиев наружу;
- регенерация;
- опорная;
- транспортная (транспорт веществ между органами)
-пищеварительная
-защитная
76. Органы осморегуляции и выделения беспозвоночных.
Органы осморегуляции и выделения беспозвоночных.
Наши беспы:
Тип Porifera — губки
Тип Placozoa — пластинчатые
Тип Ctenophora — гребневики
Тип Cnidaria — стрекающие
Тип Nematoda — нематоды
Тип Arthropoda — членистоногие
Тип Plathelminthes — плоские черви
Тип Mollusca — моллюски
Тип Annelida — кольчатые черви
Тип Rotatoria — коловратки
Пресноводные простейшие используют осморегуляцию для удаления избытка воды и для поддержания постоянной концентрации ионов в цитоплазме. Например, амёба пиноцитоз. Осморегуляция осуществляется путем активного транспорта ионов через клеточную мембрану. Кроме того, для удаления воды и ионов используется сократительная вакуоль: большого сферического пузырька, собственно сократительной вакуоли, и множества окружающих ее мембранных пузырьков, совокупность которых называется спонгиомом. Спонгиом собирает жидкость из цитоплазмы и доставляет ее в сократительную вакуоль. Вакуоль выбрасывает жидкость через пору.
Paramecium caudatum, которая живет в пресной воде, может совершать весь цикл наполнения и опорожнения вакуоли каждые 6 с.
Многие инфузории имеют по несколько вакуолей. У Paramecium по одной вакуоли имеется на обоих концах тела.
Обычно морские виды лишены сократительных вакуолей;
Аммиак, продукт выделения книдарий, хорошо растворим в воде. Он легко диффундирует через стенку тела и уносится течением. Поступающая по осмотическому градиенту вода увеличивает давление внутри полости, которая в этом случае выполняет функции гидроскелета. Избыток воды выводится через оскулюм.
|
|
|
В результате осмоса вода поступает из внешней среды в организм, избыточную воду необходимо удалить. Двусторонне-симметричные животные откачивают воду с помощью ресничных канальцев — нефридиев. В удаляемую нефридиями воду также поступают побочные продукты метаболизма. В результате образуется моча. У билатерии есть несколько видов нефридиев — фильтрационные, секреторные и почки накопления.
В фильтрационные нефридиях: образуется первичная моча — фильтрат крови, свободный от белков и клеток. Первичная моча поступает в целом, откуда откачивается парными ресничными канальцами. Эти канальцы, называемые метанефридиями, открывают ся в целом ресничной воронкой — нефростомом. Первичная моча → реабсорбция→ вторичная моча. Эти процессы осуществляются главным образом в метанефридий, но реабсорбция частично может происходить и в целоме. Реабсорбция — изъятие полезных метаболитов.
Хотя выделительные органы небольших двусторонне-симметричных животных функциональны и являются фильтрационными нефридиями, морфологически они заметно отли чаются от метанефридиальной сис темы и называются протонефридиями. Протонефридий – собой ресничный выделительный каналец, который одним концом открывается наружу, а на другом слепо замкнут жгутиковой терминальной клеткой. Терминальная клетка напоминает подоцит. Протонефридий есть у животных, лишенных кровеносных сосудов или целома. Протонефридий может включать одну или несколько терминальных клеток, каждая из которых несет один или несколько жгутиков.
Когда жгутиков много, их согласованное биение напоминает мерцающее пламя свечи. Терминальные клетки с одним жгутиком называются соленоцитами. Терминальные клетки, несущие несколько жгутиков – это мерцательные клетки (клетки с ресничным пламенем). Характерны для коловраток. Терминальные клетки этого типа встречаются у многих плоских червей.
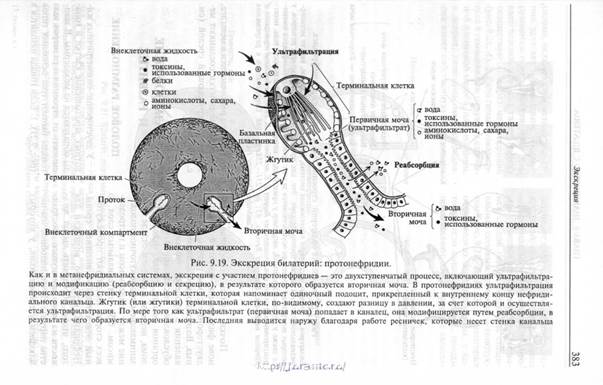
|
|
|
Азот, образовавшийся в результате метаболизма белков, турбеллярий выводят из организма в виде аммиака, который диффундирует через поверхность тела. Однако избыток воды и, вероятно, некоторые продукты метаболизма выводятся протонефридиями, несущими полицилиарные терминальные клетки.
У моллюска есть пара метанефридиев (= почек), одним концом (нефростомом) открывающихся в перикардиальную полость (целом), а другим концом (нефропором) — в выводную камеру мантийной полости. У большинства моллюсков метанефридии представляет собой мешочек с толстыми стенками, способными к секреции и абсорбции; он омывается кровью. Метанефридий → ренонерикардиальный канал → отверстие, ведущее в перикардиальную полость=нефростом → перикардиальная полость. В перикардиальной полости образуется первичная моча. Первичная моча поступает через нефростом в реноперикардиальный канал и течет по нему к нефридию. Эпителий нефридия реабсорбирует полезные вещества из мочи и возвращает их в гемолимфу. Вторичная, модифицированная моча выводится из нефридия через нефридиопор.
Сердце–почки моллюсков. 2 нефридия (почек) → 2 реноперикардиальных канала, высланные ресничным эпителием → нефростом → перикардиальная полость. Каждый нефридий открывается в выводную камеру мантийной полости нефридиопором. Гемолимфа попадает в перикардиальные железы (выпячивания перикарда, впадающие в предсердия. В железах осуществляется ультрафильтрация, и первичная моча поступает в перикардиальную полость) из предсердий. Здесь через стенку перикарда осуществляется ультрафильтрация. Первичная моча в перикард. полости → полость нефридия. → абсорбция → вторичная моча через нефридиопор в выводную камеру мантийной полости.
Экскреторные органы кольчатых червей представлены сегментарными нефридиями: метанефридиями или протонефридиями. Их свободные концы (воронки или терминальные клетки соответственно) часто располагаются на передней поверхности каждой септы. Они выступают в полость целома и омываются целомической жидкостью. Пронизывая септу, нефридиальный проток проходит в полость следующего сегмента и затем открывается во внешнюю среду. Хлорагогенная ткань, целомоциты и стенка кишечника также могут участвовать в процессах экскреции, но играют при этом второстепенную роль. Полихеты имеют или протонефридии, или метанефридиальные системы. При наличии кровеносных сосудов выделительная система полихет представлена метанефридиями; у форм, не имеющих кровеносной системы, экскреция осуществляется протонефридиями.
|
|
|
Расположенный перед септой конец метанефридия Nereis несет покрытый ресничками воронкообразный нефростом, проходящий сквозь септу в следующий сегмент каналец. Нефридиопор открывается у основания невроподии на брюшной стороне тела. По всей длине каналец выстлан ресничным эпителием.
Протонефридии полихет обычно несут группу терминальных клеток — соленоцитов. Каждая терминальная клетка имеет всего один жгутик, окруженный воротничком микроворсинок, и очень напоминает хоаноциты губок.
77. Процессы регенерации и их особенности у многоклеточных.
Способность к регенерации позволяют организму перенести утрату части клеток.
Регенерация в случае повреждения или утраты какого-нибудь органа или части организма, называется репаративной (аутотомия – отбрасывание хвоста ящерицы). Регенерацию в процессе нормальной жизнедеятельности организма называют физиологической.
В мезохиле губок: археоциты — крупные амебоидные клетки, тотипотентны, они могут дифференцироваться в клетки любого другого типа, встречающиеся у губок. Примечательна способность к регенерации у губок. Если поместить отдельные клетки в воду, то спустя некоторое время они начинают постепенно сближаться и воссоединяются, образуя целую губку, сходную с прежней. Из других животных к восстановлению целого организма из взвеси клеток способна только гидра.
Книдарии: интерстициальные клетки.
Круглые черви и коловратки, практически не способны к регенерации.
У некоторых животных со строго детерминированным развитием (эутелией) (гребневики, полихеты) во взрослом состоянии регенерация развита хорошо.
Турбеллярии: необласты – тотипотентные клетки, которые играют важную роль в заживлении ран и регенерации.
Если у планарий рассечь голову на две половинки, то рана затянется и у животного восстановятся две головы. Если планарию разрезать поперек или вдоль — каждая из половин восстановит отсутствующую часть. Регенерация головы у фрагментов, полученных из переднего конца тела червя, происходит быстрее, чем у фрагментов задней половины. Для получения нормального организма в результате регенерации достаточно всего трехсотой доли тела планарий. После того как на теле планарий сделан надрез или она разрезана пополам, эпидермис прилегающих к ране участков распространяется на ее поверхность и затягивает ее. Затем под ним образуется масса необластов, называемая бластемой.
У многих видов дождевых червей только из передней половины тела может полностью регенерировать новая особь, тогда как пиявки не способны восстановить даже отдельные утраченные органы.
78. Внутриклеточное, внекишечное и полостное пищеварение.
Внутриклеточное пищеварение характерно для представителей разных групп многоклеточных животных: губок, ресничных червей — турбеллярий, кишечнополостных, моллюсков, полихет
пищеварение происходит на основе процессов пино — и фагоцитоза. внутриклеточное пищеварение — это древний способ переваривания пищи, основанный на способности клеток к захвату пищевых частиц из внешней среды путем эндоцитоза.
Внутриклеточное пищеварение свойственно всем одноклеточным и некоторым многоклеточным организмам, находящимся па низких стадиях эволюционного развития. Оно состоит в том, что клетка (или группа клеток) активно захватывает пищевые вещества из окружающей внешней среды и при помощи специальных соков переваривает их, после чего они усваиваются цитоплазмой. В качестве примера можно привести пищеварение у амебы, которая псевдоподиями захватывает пищевые вещества и расщепляет их на простые химические соединения при помощи ферментов, находящихся в клетке. У более дифференцированных одноклеточных, как, например, у парамеции, пищевые вещества из внешней среды поступают в клетку через специальный участок цитоплазмы. В клетке вокруг пищи образуется вакуоль, где и происходит переваривание. Продукты, образовавшиеся в результате этого процесса, частично ассимилируются цитоплазмой, а частично выбрасываются наружу в виде отбросов.
Однако внутриклеточное пищеварение имеет небольшую скорость, и поэтому объем транспортируемых в клетку веществ невелик. В процессе эволюции размеры организмов увеличивались, и внутриклеточное пищеварение уже не справлялось с потребностями животных в веществе и энергии.
Эти проблемы были решены, когда возникло полостное пищеварение
Переваривание пищи происходит вне клеток, такой способ пищеварения преобладает у кольчатых червей, ракообразных, насекомых, головоногих моллюсков, асцидий, всех позвоночных животных (рыб, амфибий, пресмыкающихся, птиц, млекопитающих), в том числе и у человека.
Фактически полостное пищеварение состоит из двух этапов.
Первый этап (внеклеточное полостное пищеварение) происходит непосредственно в полости кишечника с помощью пищеварительных ферментов, секретируемых туда клетками эпителия или специальными пищеварительными железами (железами желудка, поджелудочной железой). Это происходит на относительно большом расстоянии от всасывающих клеток, поэтому такое пищеварение иногда называют внеклеточным дистантным пищеварением.
Второй этап полостного пищеварения (мембранное, или пристеночное пищеварение) происходит в непосредственной близости от всасывающих клеток кишечного эпителия на мембранах микроворсинок. Этот важный этап переваривания пищи был впервые обнаружен и подробно исследован выдающимся российским ученым А. М. Уголе- вым в конце 1950-х гг.
Мембранное пищеварение обеспечивает сложный комплекс пищеварительных ферментов, встроенных в гликокаликс мембран многочисленных апикальных микроворсинок всасывающих клеток кишечного эпителия (рис. 9. 7).
Значение этапа мембранного пищеварения очень велико:
У нематод — паразитов растений наблюдается внекишечное пищеварение. При помощи стилета в ткани растения впрыскивается секрет пищеводных желез, содержащий сильно действующие ферменты, так что начальные этапы пищеварения протекают вне организма червя. В кишечник нематоды через тонкий просвет стилета поступает уже полупереваренкая разжиженная пищевая масса
+спороцисты
у гидры сочетаются внутриклеточное и полостное пищеварение.
Внеклеточное (дистантное) пищеварение[править | править код]
Этот тип пищеварения характеризуется тем, что синтезированные секреторными клетками ферменты выделяются во внеклеточную среду, где реализуется их гидролитический эффект. Секреция гидролитических ферментов из клеток в пищеварительные полости впервые появилась у кишечнополостных и гребневиков. У кольчатых червей, ракообразных, насекомых, головоногих, оболочников и хордовых (за исключением ланцетника) внеклеточное пищеварение преобладает. Оно особенно развито у высокоорганизованных животных и человека, у которых обеспечивает начальное переваривание пищевых веществ. У этих организмов секреторные клетки расположены достаточно далеко от пищеварительных полостей, где реализуется действие гидролаз, поэтому внеклеточное пищеварение определяется как дистантное. Дистантное пищеварение, происходящее в специальных полостях, обозначается как полостное. [2]
Внеклеточное пищеварение характерно для всех гетеротрофных организмов, клетки которых имеют клеточную стенку, — бактерий, архей, грибов, хищных растений и т. д. При этом способе пищеварения ферменты секретируются во внешнюю среду или закрепляются на наружной мембране (у грамотрицательных бактерий) либо на клеточной стенке. Переваривание пищи происходит вне клетки, образовавшиеся мономеры всасываются с помощью белков-транспортеров клеточной мембраны.
Полостное пищеварение [править | править код]
Полостное пищеварение характерно для многоклеточных животных, имеющих желудочно-кишечный тракт, и происходит в полости последнего.
Внекишечное пищеварение [править | править код]
Внекишечное пищеварение характерно для некоторых животных, которые обладают кишечником, но вводят пищеварительные ферменты в тело добычи, всасывая затем полупереваренную пищу (широко известные примеры таких животных — пауки и личинки жуков-плавунцов).
Мембранное (пристеночное) пищеварение [править | править код]
Пристеночное пищеварение осуществляется в слое слизи между микроворсинками тонкого кишечника и непосредственно на их поверхности (в гликокаликсе) у позвоночных и некоторых других животных.
79. Способы оплодотворения. Преимущества внутреннего оплодотворения.
· Внутреннее оплодотворение
происходит при слиянии половых клеток (гамет) самца и самки внутри репродуктивного тракта самки. Животные, которые используют внутреннее оплодотворение, специализируются на защите развивающегося яйца. Например, рептилии и птицы откладывают оплодотворенные яйца, которые покрыты защитной оболочкой, устойчивой к потере воды и повреждениям.
· Внешнее оплодотворение
во влажных средах и требует, чтобы самец и самка выпускали или передавали свои гаметы в окружающую их среду (обычно воду). Этот процесс также называется нерестом. Преимущество внешнего оплодотворения заключается в том, что оно приводит к производству большого количества потомков. Один из недостатков заключается в том, что экологические опасности, такие как хищники, значительно уменьшают вероятность дожить до зрелого возраста. Обычно отсутствует забота о потомстве
++++++++++
· уже не требуется такое большое количество гамет (обычно все ограничивается несколькими десятками), т. к. во-первых, вероятность оплодотворения намного выше,
· зародыш защищен и его развитие не зависит от окружающей среды,
· питание зародыша осуществляется за счет материнского организма
· Большая выживаемость
80. Эмбриональное развитие. Типы личинок и их значение.
Эмбриональное развитие, как физиологический процесс, свойственно группам беспозвоночных животных у которых появляется эмбрион — стадия онтогенеза, которая проходит в яйце или матке.
Эмбриональное развитие бывает прямым или непрямым ( с метаморфозом (с превращением)). Организмы, у которых эмбриональное развитие проходит с метаморфозом, имеют личинку – ювенальная стадия, зачастую морфофизиологически отличающаяся от взрослой особи, не способна к половому размножению (за исключением неотении (способность к половому разможению у личинок), которая среди беспозвоночных животных встречается у некоторых червей и типа Arthropoda, а также педогенеза (партеногенез у личиночной стадии, встречается у типа Ctenophora и класса Trematoda)).
Яйцеклетки различаются у разных групп животных по количеству желтка, в связи с чем различаются типы дробления (ранние митотические деления зиготы, образовавшиеся в процессе дробления клетки — бластомеры).
1) Микролецитальные яйцеклетки — мелкие клетки, мало желтка, равномерно распределен, для них характерно голобластическое (по меридиальной плоскости (от анимального к вегетативному полюсам = от орального (тут прорывается бластопор) к аборальному (тут бластопора нет)) равномерное (бластомеры равных размеров) дробление. (большинство червей, представители типов Mollusca и Echinodermata (иглокожие))
2) Мезолецитальные яйцеклетки — клетки средних размеров, с умеренным количеством желтка, желток локализован на аборальном (вегетативном) полюсе. В связи с этим дробление голобластическое неравномерное. На оральном полюсе образуются мелкие микромеры, а на аборальном крупные макромеры, содержащие желток. Среди беспозвоночных представителей нет.
3) Макролецитальные яйцеклетки — крупные клетки, с большим количеством желтка, расположенным по всему объёму клетки. Дробление меробластическое — осуществляется на определенном участке клетки, борозды дробления не рассекают всю клетку, благодаря чему образуется «шапочка» из бластомеров, под которыми находится желток, бластомеры питаются диффузно. Такой тип дробление встречается у отрядов Teuthida (кальмары), Octopoda (осьминоги) среди беспозвоночных. Другой тип дробления макролецитальных яйцеклеток — апикальный (поверхностный), встречается у типа Arthropoda, в частности у класса Insecta (насекомые). Но мы их еще не знаем!
4) Алецитальные яйцеклетки — не содержат желтка, крупные, дробление голобластическое равномерное. Из беспозвоночных — тип Onychophora (онихофоры).
У трехслойных выделяют радиальное дробление (оси дробления параллельны или ортогональны анимально-вегетативной оси, Стрекающие, Лофофоры, Вторичноротые), спиральное дробление (оси дробления расположены под углом к анимально-вегетативной оси, Spiralia (плоские черви, членистоногие, кольчатые черви, моллюски и т. д. )), билатеральное дробление (билатеральная симметрия закладывается на этапе дробления, Круглые черви).
После процесса дробления формируется шар, состоящий из бластомеров, эта стадия носит название бластула:
1) Целобластула — имеет полость (Echinodermata)
2) Стерробластула — полость крайне слабо выражена, фактически отсутствует (представители группы Coelenterata, типа Mollusca, червей)
3) Плакула — двуслойная пластинка с полостью между слоями (некоторые черви)
4) Стомобластула — имеет отверстие: фиалопор (класс Calcarea)
5) Морула — полость отсутствует (некоторые представители типа Cnidaria (стрекающие))
6) Амфибластула — полость находится на анимальном полюсе, покрыта жгутиками (класс Calcarea (известковые губки))
7) Перибластула — конечная стадия апикального дробления (Insecta)
Полость бластулы — бластоцель. \
Следующая стадия онтогенеза — гаструла — двуслойный зародыш, состоящий из зародышевых листков (наружный эктодерма и внутренний энтодерма). Имеется бластопор — первичный рот, ведущий в архентерон — первичную кишку, полость которого называется гастроцель. Таксоны беспозвоночных различаются по типам гаструляции (образование гаструлы):
1) Инвагинация — впячивание клеток бластулы в бластоцель (большинство беспозвоночных)
2) Деламинация — расслоение клеток бластулы на два слоя (у некоторых Cnidaria)
3) Иммиграция — перемещение части клеток в бластоцель (у некоторых Porifera, Coelenterata, Echinodermata)
4) Эпиболия — обрастание группы клеток другими клетками с образованием двух слоев (некоторые Mollusca, Ctenophora).
Бластопор также может иметь различную судьбу:
1) Первичноротые (Protostomia) — на месте бластопора образуется рот (или и рот, и анальное отверстие). Представители: типы Брюхоресничные черви, Круглые черви, Волосатики, Морские стрелки, Моллюски, Сипункулиды, Немертины, Плоские черви, Гнатостомулиды, Коловратки, Скребни, Внутрипорошицевые, Циклиофоры, Кольчатые черви, Тихоходки, Членистоногие, Онихофоры, Мшанки, Плеченогие и Форониды. Классы Лорициферы, Киноринхи и Приапулиды. Подкласс Эхиуриды.
2) Вторичноротые (Deuterostomia) — на месте бластопора образуется анальное отверстие. Представители: типы Полухордовые, Иглокожие и Хордовые.
У трехслойных формируется мезодерма — третий зародышевый листок, данный процесс может различаться среди таксонов (например образование эпидермальной мезодермы (энтероцельный тип, засчет впячиваний архентерона (Вторичноротые)), мезенхимы (подвижные клетки, находящиеся в бластоцели) (нецеломические Spiralia) или же мезодермальные тяжи (телобластический тип, целомические Spiralia)).
Далее образуется непосредственно личинка. Личинки бывают двух типов:
1) Лецитотрофные — питаются желтком
2) Планктотрофные — питаются планктоном
Основное значение личинок: расселение, нахождение благоприятного места для жизнедеятельности взрослой особи (личинка должна осесть), проникновение в хозяина (у паразитов).
81. Эволюция кожно-мускульного мешка беспозвоночных животных.
У турбелярий Наиболее поверхностно располагается слой кольцевых мышц, несколько глубже — продольных и, наконец, диагональных мышечных волокон, идущих в двух направлениях и перекрещивающихся между собой под углом. Совокупность мышц вместе с эпителием образует кожно-мускульный мешок. Кроме того, у всех турбеллярий имеются характерные для них дорзовентральные, или спиннобрюшные, мышцы НЕ ВХОДЯТ В КММ.
КДП
У сосальщиков КП тегумент
У ленточных КП Непосредственно под базальной мембраной располагается наружный слой кольцевых и внутренний — продольных мускульных волокон. Часто к этим двум слоям присоединяется еще третий более глубокий слой кольцевых мышечных волокон (рис. 156).
Под гиподермой лежит мускулатура, которая у круглых состоит лишь из одного слоя, а именно — продольных волокон. Мускульный слой несплошной, но разбит валиками гиподермы на 4 продольные ленты. При сокращении спинные и брюшные ленты действуют как мышцы антагонисты, и тело нематод способно изгибаться только в дорзовентральной плоскости. При этом червь, как правило, передвигается на боку. Таким образом, функционально брюшной стороной будет одна из боковых сторон нематоды — правая или левая. Строение мышечных волокон крайне своеобразно. Каждая мышечная клетка имеет форму длинного веретена (например, у лошадиной аскариды достигает 0, 5 см длины), от центра которого на стороне, обращенной к полости тела, выдается большой пузыревидный придаток (рис. 179). Периферия волокна содержит тончайшие сократительные фибриллы, тогда как середина и пузыревидный придаток состоят из цитоплазмы; в придатке помещается и ядро мышечной клетки.
Кожно-мускульный мешок у коловраток отсутствует. В теле имеются отдельные, большей частью поперечнополосатые мышечные волокна, служащие для втягивания головного отдела, для сокращения туловища и т. д. Непосредственно под наружным эпителием лежит первичная полость тела.
Под кожным эпителием полихет находится хорошо развитый двухслойный мускульный мешок, состоящий из наружных кольцевых и внутренних продольных мышц (рис. 216). Продольная мускулатура у примитивных форм образует сплошной слой, у прочих же разбивается основаниями параподий на 4 продольные ленты: две из них лежа т ближе к брюшной, две — ближе к спинной стороне. Внутренняя сторона продольного слоя мышц выстлана однослойным перитонеальным эпителием мезодермального происхождения, под ним находится обширная вторичная полость тела, или целом
У олигохет хорошо развитые слои кожно-мускульного мешка — наружный кольцевой и более мощный внутренний продольный. Обширный целом поделен хорошо развитыми диссепиментами и брюшным мезентерием, спинной мезентерий'отсутствует
Тело одето снаружи довольно плотной кутикулой; лежащий под ней эпителий очень богат слизистыми железистыми клетками, а у основания эпителиальных клеток рассеяны многочисленные пигментные клетки с зернистостью различного цвета. Присутствие этих клеток обусловливает окраску пиявок. Под эпителием располагаются кольцевые и очень сильно развитые продольные мышцы,
82. Органеллы и органы активной защиты и нападения.
У инфузорий экструсомы (трихоцисты выствреливают вещество, застывающее в нити; токсицисты – выд токс в-ва; мукоцисты- выд слизь удерживают добычу, образ слизистый домик для цисты; гаптоцисты – заякоривание и контакта с пищей) Цирры – кисточки из ресничек для бега шага прыжков.
У кишечнополостных стрекательные клетки – книдоциты пенетрант- гарпун, глютинант-липкая нить, вольвент-обкручивает жертву тонкая нить. Щупальца
6. Щупальца(поражение и удерживание добычи(Медузы, Гребневики(клейкие шупальца)) 7. Неодермис(тегумент)-защитный покров (Класс Сосальщики)
8. Многослойная кутикула(прочность и эластичность покровов)-(Тип Круглые черви) 9. Коловращательный аппарат(приток воды с пищей)
10. Глотка-стилет, пробуравливающая покровы тела добычи(Хоботные пиявки)
11. Глотка с челюстями-прокусывание покровов добычи(Челюстные пиявки)
12. Лигамент-удерживает створки раковин закрытыми(Двустворчатые моллюски)
13. Чернильный мешок с грушевидной железой(Головоногие)
ля овладения добычей и её поедания головоногим служат многочисленные присоски, которыми усеяны с внутренней стороны их мускулистые щупальца, и челюстной аппарат в виде крепкого клюва, по форме похожего на клюв попугаев.
Прежде всего его окраска становится более тёмной и ещё более приметной для хищника, а затем он выбрасывает из чернильного мешка его содержимое, но не в виде расплывающейся в воде тёмной краски, а в форме тёмного тела, в котором чернильная жидкость заключена в тонкостенную плёнку и которое в таком виде представляет собой грубое подобие тела самого кальмара.
Таким образом преследуемый моллюск как бы подменяет себя своим макетом, или «чернильным чучелом», на которое и устремляется хищник, в то время как сам моллюск быстро принимает бледную окраску и уходит из поля зрения преследователя. Хищник легко настигает подброшенное ему «чучело» кальмара и разрушает одевавшую его тончайшую плёнку; чернильная жидкость расплывается в воде и при этом не только скрывает от глаз преследователя ускользнувшую от него добычу, но содержащимся в ней наркотическим веществом лишает его чувства обоняния; ясно, что при таких условиях дальнейшее преследование ускользнувшего головоногого становится уже невозможным: моллюск спасён.
У многих видов современных головоногих встречается и ряд других защитных приспособлений: светящиеся органы, выделение светящейся слизи, а у молоди одного средиземноморского осьминога жгучие обрывки медуз используются в качестве защитного оружия.
У хоботных пиявок (отр. КЬупсЬоЬс1е1Нс1а) глотка образует мускулистую трубку, или хоботок, способную временно высовываться из рта для на падения на добычу. У челюстных (отр. ОпаШоМеПЫа) глотка не выворачивается, но в ротовой полости имеются три мускулистых валика, 1 спинной и 2 боковых (рис. 238). По свободному краю каждого валика сидит ряд хитиновых зубчиков, совокупность которых образует зазубренную вроде пилы челюсть. Во время принятия пищи челюсти пиявки прорезают кожу хозяина: получается трехлучевая ранка, из которой пиявка и высасывает кровь.
При помощи биссусовой нити и шиповатых створок глохидии прикрепляются к жабрам или плавникам рыбы. Вокруг вызванной глохидиями маленькой ранки раздражаемый эпителий рыбы разрастается и постепенно покрывает личинку. Внутри получившейся опу холи глохидий питается за счет хозяина, увеличивается в размерах и в течение нескольких недель проходит дальнейшее развитие.
83. Органы газообмена у беспозвоночных животных.
· Моллюски: первичные жабры (ктенидии), вторичные жабры, легкие.
· Плоские и круглые черви: дыхательная система отсутствует.
· Кишечнополостные: дыхательная система отсутствует.
· Губки: дыхательная система отсутствует.
· Пластинчатые: дыхательная система отсутствует
Органы дыхания многощетинковых довольно разнообразны. Наиболее просто организованные представители дышат всей поверхностью тела подобно низшим червям. У большинства функцию дыхания принимают па себя известные участки параподий. Часто спинной усик превращается в жабру (см. рис. 215, В). Внутрь ее заходят кровеносные сосуды, а через стенки растворенный в воде кислород поступает в кровь. Жабры имеют листовидную, перистую или кустистую форму и развиваются большей частью не на всем протяжении тела, а только на известном его участке.
Панцирные моллюски обладают жабрами. Число их в отличие от большинства моллюсков колеблется от 4 до 80 пар. Жабры располагаются в один ряд на каждой стороне тела в мантийной борозде (см. рис. 425), причем одна пара, лежащая позади почечных отверстий, больше других. Каждая жабра имеет вид ланцетовидной пластинки с широким основанием; на обеих плоских сторонах пластинки перпендикулярно к ней отходит по ряду лепестков, прилегающих друг к другу, как страницы книги. Таким образом, жабра имеет двоякоперистое строение. Вся поверхность ее одета мерцательным эпителием. Вдоль узкой стороны жаберной пластинки, обращенной к ноге, от основания к свободному концу жабры проходит приносящий жаберный сосуд с венозной кровью, а вдоль стороны, обращенной к мантии, от конца жабры к ее основанию идет выносящий сосуд с окисленной кровью. Веточки обоих сосудов продолжаются в жаберные лепестки. Благодаря работе ресничек эпителия вокруг жабры циркулирует вода, что способствует газообмену между кровью жаберных сосудов и внешней средой.
Большинство брюхоногих дышит жабрами. Первичными, или настоящими, жабрами являются ктенидии, парные органы, расположенные но бокам от порошицы. У многих форм они имеют вид вытянутых двоякоперистых придатков, заостряющихся к свободному концу. Каждый ктенидий состоит из осевого уплощенного стволика, несущего два ряда лепестков. Ктенидии характеризуются присутствием у их основания органов химического чувства — осфрадиев. Первично ктснидиев одна пара (рис. 447), но в связи с недоразвитием органов правой стороны тела у большинства брюхоногих правый ктенидий атрофируется. Так, у некоторых переднежаберных (НаИоШ) он меньше левого. У высших представителей этой группы жабра уже одна, и притом нередко из двоякоперистой она становится однорядно
|
|
|
