 |


Неодарвинистская теория эволюции
|
|
|
|
Фактически очень мало известно, если вообще когда-нибудь может быть известно, о деталях эволюции в прошлом. Также эволюцию нелегко наблюдать в настоящем. Даже на шкале времени в миллионы лет начало новых видов является редким, а родов, семейств и отрядов — еще более редким событием. Эволюционные изменения, которые действительно наблюдались в течение последнего столетия или около того, большей частью касаются развития новых разновидностей или сортов в пределах установленных видов. Примером, наиболее подтвержденным документально, может служить появление разновидностей нескольких европейских мотыльков с темной окраской в районах, где промышленные выбросы привели к почернению поверхностей, на которых они обитают. Темные мутанты лучше выживали вследствие естественного отбора, поскольку были лучше замаскированы и потому их меньше поедали птицы.
При таком скудном наборе прямых свидетельств и при столь малых возможностях экспериментальной проверки любая интерпретация механизма эволюции неизбежно должна быть спекулятивной: не ограниченная конкретными фактами, она будет представлять собой главным образом различные варианты развития своих изначальных предположений о природе наследственности и об источниках наследственной изменчивости.
Ортодоксальная механистическая интерпретация дается неодарвинистской теорией, которая отличается от оригинальной теории Дарвина по двум основным моментам: во-первых, она предполагает, что наследственность может быть объяснена с помощью генов и хромосом; и во-вторых, что конечной причиной наследственной изменчивости являются случайные мутации генетического материала. Основные положения этой теории можно суммировать следующим образом:
|
|
|
(1) Мутации происходят случайно.
(2) Гены рекомбинируют путем полового воспроизведения, перекрещивания (кроссинговера) хромосом и изменения их структуры. Эти процессы производят новые перестановки (пермутации) генов, которые могут вызывать новые эффекты.
(3) Распространение благоприятной мутации протекает быстрее в малых, нежели в больших скрещивающихся, популяциях. В малых или среднего размера популяциях, которые претерпевают большие флуктуации, мутантные гены могут быть утеряны или сохранены наугад скорее путем «генетического дрейфа», чем в результате естественного отбора.
(4) Естественный отбор стремится исключить мутантные гены с вредными эффектами. Факторы отбора включают: хищников, паразитов и инфекционные заболевания, конкуренцию за пространство, еду и т. д., климатические и микроклиматические условия и сексуальный отбор.
(5) Новые давления отбора выступают на сцену в результате изменений условий среды и моделей по ведения самих организмов.
(6) Если популяции разделяются географически, или экологически, или по какой-либо другой причине, то вероятно, что их эволюция становится дивергентной.
(7) В особенности в растительном царстве новые виды могут появляться от внутривидовых гибридов, которые хотя обычно стерильны, но иногда становятся плодоносящими вследствие полиплоидности.
Некоторые из основных положений этой неодарвинистской теории были разработаны математически в области теоретической популяционной генетики. Для построения математических моделей обычно принимается простоты ради, что гены подвергаются отбору независимо друг от друга (хотя на самом деле они связаны вместе в хромосомах и при своих проявлениях взаимодействуют с другими генами). Тогда, приписывая численные значения давлениям отбора, скоростям мутаций и размерам популяций, можно определить изменения частоты гена на протяжении данного числа поколений. Эти методы были распространены на все аспекты эволюции с помощью допущения, что морфологические признаки и инстинкты определяются отдельными генами или комбинациями генов[163].
|
|
|
Большинство неодарвинистских теоретиков полагают, что дивергентная эволюция под влиянием естественного отбора на протяжении длительных периодов времени приводит к развитию не только новых сортов, разновидностей и подвидов, но также новых видов, родов, семейств, отрядов и типов[164]. Этот взгляд оспаривался на том основании, что различия между этими высшими таксономическими единицами слишком велики, чтобы они могли возникать в результате постепенных трансформаций; помимо чего-либо другого организмы часто различаются количеством и структурой своих хромосом. Разные авторы высказывали мнение, что эти крупномасштабные эволюционные изменения происходят внезапно в результате макромутаций. Современными примерами таких внезапных изменений могут служить уродливые животные и растения (монстры), у которых структуры были трансформированы, дублированы или подавлены. Случайно в процессе эволюции «многообещающие монстры» могли выживать и воспроизводиться[165]. Один из аргументов в пользу такого взгляда состоит в том, что, в то время как постепенные изменения под давлением отбора должны приводить к формам с различной адаптивной ценностью (за исключением, быть может, малых популяций, подверженных «генетическому дрейфу»), макромутации могут производить все типы кажущихся беспричинными макромасштабных вариаций, которые могут быть удалены путем естественного отбора, только если они несомненно вредны. Это помогает объяснить удивительное разнообразие живых организмов[166].
Несмотря на то что эти авторы подчеркивают важность внезапных больших изменений, они не отвергают также ортодоксальные положения о том, что эволюция в целом зависит только от случайных мутаций и генетической наследственности в сочетании с естественным отбором.
Более радикально настроенные критики оспаривают сами эти фундаментальные принципы, утверждая, что вряд ли можно допустить, что все адаптивные структуры и инстинкты живых организмов могли возникнуть чисто случайно, даже если полагать, что естественный отбор позволяет организмам выживать и воспроизводиться, только если они достаточно хорошо к этому приспособлены. Более того, они утверждают, что некоторые случаи параллельной и конвергентной эволюции, в которых очень сходные морфологические признаки появляются независимо в различных таксономических группах, указывают на действие в эволюции неизвестных факторов, даже если допустить существование параллельно действующих давлений отбора. Наконец, некоторые возражают против явно или неявно выраженного механистического положения, что эволюция в целом совершенно бесцельна[167].
|
|
|
Метафизическое отрицание какой-либо творческой силы или цели в эволюционном процессе вытекает из материалистической философии, с которой механистическая теория столь тесно связана[168]. Но если не считать, что результаты научных и метафизических исследований должны быть безнадежно перепутаны, в рамках ограниченного контекста эмпирической науки неодарвинизм должен рассматриваться не как метафизическая догма, но как научная гипотеза. А как таковая она вряд ли может считаться доказанной: в лучшем случае она предлагает вероятную интерпретацию процессов эволюции на основе своего предположения о генетической наследственности и случайности мутаций.
Гипотеза формативной причинности позволяет рассматривать наследственность в новом свете и поэтому приводит к существенно иной интерпретации эволюции. Но, находясь в согласии с неодарвинистским положением о случайности генетических мутаций, она не подтверждает и не отрицает метафизику материализма (раздел 8.7).
Мутации
Если бы организмы развивались поколение за поколением в одних и тех же окружающих условиях и передавали своему потомству все время одни и те же гены, сочетание эффектов генетической наследственности и морфического резонанса привело бы к бесконечному повторению одних и тех же старых форм. Но в действительности организмы претерпевают изменения под влиянием факторов как внутренних — генетических мутаций, так и внешних — изменений окружающей среды.
|
|
|
Мутации представляют собой случайные изменения структуры генов или хромосом, непредсказуемые для данного индивидуума не только на практике, но также и в принципе, поскольку они зависят от вероятностных событий. Нет причин сомневаться, что они носят случайный характер, как и предполагает неодарвинистская теория.
Многие мутации вызывают эффекты настолько вредные, что они становятся смертельными. Но из тех, которые менее опасны, некоторые влияют на морфогенез посредством количественных воздействий на пути морфогенеза и приводят к появлению различных вариантов нормальных форм (раздел 7.3), а другие влияют на морфогенетические зародыши таким образом, что все пути морфогенеза либо полностью блокируются, либо заменяются на иные (раздел 7.2).
В тех редких случаях, когда мутации ведут к изменениям, которым благоприятствует естественный отбор, не только возрастает доля мутантных генов в популяции, в согласии с неодарвинистской теорией, но также повторение новых путей морфогенеза в увеличивающемся количестве особей будет усиливать новые хреоды; не только «пулы генов», но и морфогенетические поля вида будут изменяться и эволюционировать вследствие естественного отбора.
Дивергенция хреод
Если мутация или изменение окружения нарушает нормальный ход морфогенеза на сравнительно ранней стадии, система может быть способна к регулированию и продолжению развития до создания нормальной формы, несмотря на это нарушение. Если этот процесс повторяется поколение за поколением, отклонение хреоды будет стабилизироваться морфическим резонансом; следовательно, вся порода или разновидность данного вида начнет следовать ненормальному пути морфогенеза, хотя в результате получится обычная взрослая форма. Действительно, описано много случаев так называемых временных отклонений в развитии. Например, у торфяных червей Prorhynchnus stagnitilis яйцеклетки делятся либо спиральным, либо радиальным способом, и развивающиеся эмбрионы растут либо внутри желтка, либо на его поверхности. Вследствие этих различий на ранних стадиях эмбрионального развития некоторые органы формируются в различной последовательности; тем не менее взрослые животные идентичны. А у единственного вида кольчатого червя, Nereis, образуются два очень разных типа личинок; несмотря на это, обе развиваются в одну и ту же взрослую форму[169]. В некоторых из таких случаев временные отклонения могут быть адаптивными, например, к условиям жизни личинок, но по большей части они возникают без видимых причин. Гораздо большее эволюционное значение имеют те отклонения хреод, которые не полностью корректируются путем регуляции и поэтому приводят к появлению отличающихся друг от друга конечных форм. Такие изменения путей развития могут возникать в результате мутаций (ср. раздел 7.3) или необычных условий окружающей среды (ср. раздел 7.6). В случае мутации в неизменном окружении, если отклонившаяся конечная форма имеет преимущество с точки зрения отбора, частота мутантных генов в популяции будет возрастать и, кроме того, новая хреода усилится вследствие морфического резонанса. В более сложном случае, когда отклонившаяся форма, возникшая в ответ на необычные окружающие условия, имеет селективное преимущество, новая хреода будет усиливаться, как и раньше, и в то же время отбор будет действовать в пользу организмов, имеющих генетические возможности для такой реакции на окружение (сравните с экспериментами Уоддингтона на плодовых мушках, раздел 7.7). Так приобретенные признаки станут наследуемыми в результате сочетания генетического отбора и морфического резонанса.
|
|
|
В естественных условиях действие различных давлений отбора на географически или экологически изолированные популяции какого-либо вида приведет к расхождению (дивергенции) как их «генных пулов», так и их хреод. Действительно, бесчисленные виды животных и растений разошлись (в ходе эволюции) в генетически и морфологически различные породы и разновидности; знакомыми примерами являются одомашненные животные и культурные растения[170]. Подумайте, например, о поразительно различающихся породах собак, от афганской гончей до китайского мопса.

Рис. 20. Сравнение форм различных видов рыб (из Thompson, 1942). Воспроизведено с разрешения издательства Cambridge University Press
В некоторых случаях морфологическое расхождение затрагивает лишь одну определенную структуру или малую группу структур, тогда как другие остаются относительно устойчивыми. Например, в маленькой рыбке Beloneacus на ранних стадиях развития челюсти подобны таковым у родственных видов, но впоследствии развиваются в необычайно удлиненное рыло[171]. Множество различных структурных преувеличений развились под влиянием сексуального отбора, например рога у оленя. А цветы дают тысячи примеров расхождений в развитии различных составных частей: сравните, например, модификации лепестков у разных видов орхидей.
В других случаях форма многих различных структур меняется сходным образом. Если формы варьируются особенно единообразно и гармонично, их можно сравнивать графически, по систематическому искривлению наложенных координат (рис. 20), как показал сэр Д'Арси Томпсон в разделе своего очерка «О росте и форме», озаглавленном «Теория трансформаций, или сравнение родственных форм».
Эволюционные изменения такого типа происходят в рамках уже существующих морфогенетических полей. Они создают вариации на данные темы. Но они не могут объяснить существование самих этих тем. По словам Томпсона:
«Мы не можем превратить беспозвоночное в позвоночное или кишечнополостное в червя посредством какой-либо простой и правильной деформации или чего-либо, не поддающегося разложению на элементарные принципы... Формальное сходство, от которого мы зависим как от надежного критерия родства животных в некоторых пределах или при некоторых степенях близости и подобия, в других случаях перестает нам служить, поскольку при некоторых обстоятельствах оно перестает существовать. Наши геометрические аналогии являются весомым свидетельством против дарвиновской концепции бесчисленных и непрерывных изменений; они помогают показать, что скачкообразные изменения естественны, что..внезапные изменения, большие или меньшие, должны были иметь место и время от времени должны были возникать новые типы» [172].
Подавление хреод
В то время как дивергенция хреод в пределах существующих морфогенетических полей допускает непрерывные или количественные изменения формы, изменения в развитии, влекущие за собой подавление хреод или замену одной хреоды на другую, приводят к качественным скачкам. Согласно гипотезе формативной причинности, такие эффекты вызываются мутациями или факторами среды, которые изменяют морфогенетические зародыши (раздел 7.2). Примеры мутантных листьев гороха, в которых усики заменены на листочки, показаны на рис. 18, а мутант дрозофилы с «двойной грудью» — на рис. 17.

Рис. 21. Росток вида Acacia (Gebel, 1898)
Изменения такого типа, вероятно, часто происходили в ходе эволюции. Например, у некоторых видов акации листья были подавлены и их роль играли уплощенные черенки. Этот процесс можно увидеть на ростках, у которых первые образовавшиеся листья являются типично перистыми (рис. 21). У членов семейства кактусов листья были заменены на шипы. Среди насекомых почти в каждом отряде есть виды, у которых крылья подавлены либо у обоих полов, как у некоторых паразитирующих мух, либо только у одного пола, как у самки жука, известной как личинка светляка обыкновенного (glow worm). У муравьев личинки самок развиваются либо в крылатых королев, либо в бескрылых работниц в зависимости от химического состава их пищи.
У некоторых видов подростковые формы становятся половозрелыми и воспроизводятся, не производя когда-либо структур, характерных для взрослых форм, которые, так сказать, замкнуты накоротко. Классическим примером является аксолотль, головастик тигровой саламандры, который достигает полного размера и становится сексуально зрелым, не теряя личиночных признаков. Если аксолотли получают тироидный гормон, они превращаются в дышащую воздухом взрослую форму и выходят из воды на сушу.
Самые яркие примеры подавления хреод встречаются у паразитов, среди которых есть такие, которые утеряли почти все структуры, характерные для родственных свободно живущих форм.
Повторение хреод
У всех многоклеточных организмов некоторые структуры повторяются несколько или много раз: щупальца гидры или морской звезды, ноги многоножки, перья птиц, листья деревьев и так далее. Кроме того, многие органы построены из повторяющихся структурных единиц: трубочки почек, сегменты фруктов и т. д. И конечно, на микроскопическом уровне ткани содержат тысячи или миллионы копий немногих основных типов клеток.

Рис. 22. Груша-монстр (Masters, 1869)
Если вследствие мутаций или изменений окружающей среды в развивающихся организмах образуются дополнительные морфогенетические зародыши, тогда некоторые структуры могут быть повторены большее число раз, чем обычно. Примером, знакомым садоводам, являются двойные цветки, содержащие добавочные лепестки. Иногда у человека младенцы рождаются с добавочными пальцами на руках или ногах. Много примеров ненормально умноженных структур можно найти в учебниках по тератологии — от телят с двумя головами до уродливых многоплодных груш (рис. 22).
По мере развития этих дополнительных структур регуляция происходит таким образом, что они более или менее полно объединяются с остальной частью организма: например, добавочные лепестки в двойных цветках имеют нормальные сосудистые соединения, а добавочные пальцы на руках или ногах — нормальные кровоснабжение и иннервацию.
То, что такое умножение структурных единиц должно было играть существенную роль в эволюции новых типов животных и растений, очевидно из структурных повторений у существующих организмов. Более того, многие структуры животных и растений, которые сейчас отличаются друг от друга, вполне могли развиться из изначально одинаковых единиц. Считается, например, что насекомые развились из существ, напоминающих примитивных многоножек, имеющих ряд более или менее одинаковых сегментов, каждый из которых несет одну пару ногоподобных придатков. Из этих придатков на передних сегментах могли развиться ротовая часть и щупальца или усики, а из самих сегментов, слившихся воедино,— голова. В хвостовой части некоторые из придатков могли видоизмениться с образованием структур, предназначенных для спаривания и кладки яиц. В сегментах брюшной области придатки были подавлены, но в трех грудных сегментах они сохранились и развились в ноги современного насекомого[173].
Такая дивергенция изначально сходных хреод могла стать возможной, только если морфогенетические зародыши сегментов стали различаться по своей структуре; в противном случае все они продолжали бы связываться с помощью морфического резонанса с одними и теми же морфогенетическими полями. И даже у современных насекомых, если бы на ранних стадиях их развития не происходило такого расхождения сегментальных зачатков, нормальное различие между сегментами было бы утеряно. Действительно, это именно то, что, по-видимому, происходит у плодовой мушки дрозофилы в результате мутаций в удвоенном грудном генном комплексе: некоторые из них трансформируют структуры третьего грудного сегмента в структуры второго, так что мушка имеет две пары крыльев вместо одной (рис. 17); другие трансформируют брюшные сегменты в сегменты грудного типа, несущие ноги; а есть и такие, которые вызывают противоположный эффект, превращая грудные сегменты в сегменты брюшного типа[174].
Влияние других видов
Те, кто практикует разведение животных и растений, давно заметили, что культурные разновидности время от времени дают потомство, напоминающее их диких предков. Более того, когда скрещиваются две определенные культурные разновидности, потомство иногда имеет признаки не кого-либо из родителей, а, скорее, диких предков. Этот феномен называют обращением (реверсией), или атавизмом[175].
Подобным же образом в эволюции некоторые типы морфологических отклонений можно рассматривать как реверсии путей развития более или менее отдаленных диких видов. Например, ненормальное образование двух пар крыльев в мутантах дрозофилы с «двойной грудью» {рис. 17) интерпретировалось как отбрасывание назад к типу развития, характерному для четырехкрылых предков этих мушек[176]. Множество других примеров предполагаемого атавизма можно найти в литературе по тератологии[177]. Конечно, такие интерпретации могут быть лишь спекулятивными, но они не обязательно далеки от истины. Мутации или ненормальное окружение могли создать в эмбриональных тканях условия, сходные с теми, которые были у диких предков, с теми же морфогенетическими последствиями.
У большинства растений и животных лишь малая часть, быть может менее пяти процентов, хромосомной ДНК содержит гены, кодирующие белки данного организма. Функции огромного большинства молекул ДНК остаются неизвестными. Некоторые могут участвовать в контроле синтеза белка, некоторые — играть структурную роль в хромосомах, а некоторые могут состоять из «лишних» наследованных генов, которые более не проявляются (не экспрессируются). Было высказано предположение, что если мутация — например, из-за перестановки в структуре хромосомы — приводит к экспрессии таких латентных генов, неожиданно снова могут начать синтезироваться белки, характерные для далеких предков, в результате чего в некоторых случаях вновь появляются давно утерянные структуры[178].
В рамках гипотезы формативной причинности, если какое-либо из таких изменений являлось причиной того, что морфогенетический зародыш принимал структуру и вибрационную картину, сходные с теми, которые были у какого-либо вида этого рода, он подпадает под влияние морфогенетического поля вида, даже если он вымер миллионы лет назад. Более того, этот эффект необязательно ограничен родовыми типами. Если в результате мутации (или по какой-нибудь другой причине) структура зародыша в развивающемся организме стала достаточно близкой к таковой у морфогенетического зародыша любого другого вида, современного или вымершего, он будет «настроен» на хреоду, характерную для этого другого вида. И если клетки способны синтезировать соответствующие белки, тогда система фактически будет развиваться под его влиянием.
В ходе эволюции очень близкие структуры иногда появляются как бы совершенно независимо в линиях, состоящих в более или менее отдаленном родстве. Например, среди средиземноморских сухопутных улиток виды, относящиеся к хорошо различимым родам, определяемым по их гениталиям, имеют раковины почти одинаковой формы и структуры; рода ископаемых аммонитов демонстрируют повторяющееся параллельное развитие раковин с килем и желобками, а подобный или идентичный рисунок крыльев встречается в совершенно разных семействах бабочек[179].
Если мутация привела к тому, что организм «настроился» на хреоды других видов и, следовательно, в нем развиваются структуры, характерные для других видов, он вскоре будет уничтожен в ходе естественного отбора, если эти структуры уменьшают его шансы на выживание. С другой стороны, если он будет «поощряться» естественным отбором, доля таких организмов в популяции будет увеличиваться. Действительно, давления отбора, которые способствовали увеличению, могут сильно напоминать те, которые способствовали начальной эволюции этого особого признака у других видов. А иногда структурное сходство может поддерживаться даже просто само по себе, именно потому что оно позволяет организму копировать особей других видов. Таким образом, эволюционные параллелизмы могут часто зависеть как от «подбирания» одним видом морфогенетических полей другого, так и от параллельных давлений отбора.
С другой стороны, сходные давления отбора могут приводить также к конвергентной эволюции избыточных подобных структур у различных видов посредством модификации разных морфогенетических полей. Но в таких случаях, если структуры не сильно похожи друг на друга во внутренних деталях и по внешней форме, они вряд ли могут взаимодействовать через морфический резонанс.
Источник новых форм
Согласно гипотезе формативной причинности, морфический резонанс и генетическая наследственность в совокупности объясняют повторение характерных путей морфогенеза в последовательных поколениях растений и животных. Более того, признаки, приобретенные в ответ на влияния окружающей среды, могут стать наследуемыми в результате сочетания морфического резонанса и генетического отбора. Морфология организмов может изменяться вследствие подавления или повторения хреод; и некоторые поразительные примеры параллельной эволюции можно объяснить «переходом» хреод от одного вида к другому.
Однако ни повторение, модификация, добавление или вычитание, ни перестановка существующих морфогенетических полей не могут объяснить источник самих этих полей. Тем не менее в процессе эволюции должны были появляться совершенно новые морфические единицы вместе со своими морфогенетическими полями: это поля органелл — основных типов клеток, тканей и органов, а также поля фундаментально различающихся типов низших и высших растений и животных.
Хотя генетические мутации и ненормальное окружение вполне могли предоставить случаи для первого появления новых биологических морфических единиц, формы их морфогенетических полей не могли быть полностью определены ни энергетической причинностью, ни ранее существовавшими формативными причинами (раздел 5.1). Можно лишь высказывать догадки о том, появилось ли любое данное морфогенетическое поле внезапно, в виде большого «скачка», или более постепенно, в последовательности меньших «скачков». Но в любом случае новые формы, принимавшиеся в этих «скачках», не могут быть объяснены в рамках научного подхода через предшествующие причины.
Происхождение новых форм можно приписать либо творческой активности силы, наполняющей собой природу и трансцендентной ей; либо творческому импульсу, присущему природе; либо слепому и бесцельному случаю. Но выбор между этими метафизическими возможностями никогда не мог быть сделан на основе какой-либо научной гипотезы, проверяемой опытным путем. Поэтому с точки зрения естественной науки вопрос об эволюционном творчестве можно лишь оставить открытым.
Глава 9
Движения и моторные поля
Введение
В предыдущих главах обсуждалась роль формативной причинности в морфогенезе. В этой и двух последующих главах речь пойдет о роли формативной причинности в управлении движением.
Некоторые движения растений и животных являются самопроизвольными (спонтанными); это означает, что они происходят без какого-либо определенного стимула извне. Другие движения совершаются в ответ на стимул из среды. Конечно, организмы пассивно отвечают на действие грубой физической силы: дерево может быть повалено ветром или животное может унести сильный поток воды,— но многие отклики активны и не могут быть объяснены как результат грубого физического или химического воздействия стимула на организм в целом: они выявляют чувствительность (сенситивность) организма к влияниям среды. Эта чувствительность обычно зависит от специализированных рецепторов или органов чувств.
Физико-химическая основа возбуждения таких специализированных рецепторов стимулом со стороны среды была выяснена весьма подробно; основательно исследованы также физиология нервного импульса, функционирование мускулов и других моторных структур. Но еще очень мало известно об управлении и координации поведения.
В этой главе предполагается, что как формативная причинность организует морфогенез через вероятностные структуры полей, которые определенным образом упорядочивают энергетически неопределенные процессы, так же она организует и движения и, следовательно, поведение. Подобие между морфогенезом и поведением сразу не очевидно, но его легче всего понять на примере растений и одноклеточных животных, таких как амеба, движения которых существенно морфогенетичны. Их мы и рассмотрим в первую очередь.
Движения растений
Обычно растения движутся путем роста[180]. Этот факт легче осознать, когда видишь их в ускоренной киносъемке: ростки вытягиваются и изгибаются к свету; разветвления корней устремляются вниз, в почву, а верхушки усиков и ползучих стеблей выбрасывают в воздух широкие спирали, пока не соприкоснутся с твердой опорой и не обовьются вокруг нее[181].
Рост и развитие растений происходят под контролем их морфогенетических полей, которые сообщают растениям их характерные формы. Но ориентация этого роста в значительной степени определяется направленными стимулами силы тяжести и света. Факторы окружения влияют также на тип развития: например, в тусклом свете растения становятся белесыми, их побеги растут сравнительно быстро и становятся длинными и тонкими, пока не достигнут более яркого света.
Гравитация «чувствуется» благодаря ее действию на зерна крахмала, которые скатываются вниз и накапливаются в нижних частях клеток[182]. Направление, откуда приходит свет, обнаруживается путем дифференциального поглощения лучистой энергии на освещенной и теневой сторонах органов желтым пигментом каротеноидом[183]. Чувство «осязания», с помощью которого ползучие стебли и усики находят твердые опоры, может объясняться выделением простого химического вещества, этилена, с поверхности клеток при их механической стимуляции[184]. Изменение роста с ускоренного при недостатке света на нормальный зависит от поглощения света голубым белковым пигментом, называемым фитохромом[185].
Ответы на эти стимулы включают сложные физико-химические изменения в клетках и тканях, а в некоторых случаях зависят от различий в распределении гормонов, таких как ауксин. Однако эти реакции не могут быть объяснены с помощью одних только физико-химических изменений, но могут быть поняты лишь с привлечением общих морфогенетических полей растений. Например, благодаря присущей им полярности растения производят на одном конце стебли, на другом — корни. Направленный стимул гравитации руководит этим поляризованным развитием так, что стебли растут вверх, а корни вниз. Действие гравитационного поля на зерна крахмала в клетках и соответствующие изменения в распределении гормонов действительно являются причиной этих ориентированных движений роста, но не могут сами по себе объяснить существование полярности; не объясняют они и тот факт, что главные стебли и корни реагируют диаметрально противоположным образом, а также различные особенности роста деревьев, трав, вьющихся и ползучих растений и особенности разветвления стебля и корневой системы различных видов. Все эти признаки зависят от морфогенетических полей.
Несмотря на то что большая часть движений растений происходит только в молодых растущих органах, некоторые структуры сохраняют способность двигаться даже тогда, когда они вполне созрели, например, цветы, которые открываются и закрываются каждый день, и листья, складывающиеся ночью. На эти движения влияют интенсивность света и другие факторы среды; они находятся также под контролем «физиологических часов» и продолжают происходить приблизительно раз в сутки, даже если растения помещаются в неизменяемые условия[186]. Листья или лепестки открываются, потому что специализированные клетки в «шарнирной» (hinge) области у их основания разбухают; и они закрываются, когда эти клетки теряют воду вследствие изменения проницаемости их мембран по отношению к неорганическим ионам[187]. Восстановление тургорного давления является активным процессом, требующим затрат энергии и сравнимым с ростом.
Помимо «сонных» движений листья некоторых растений движутся в течение дня в ответ на изменение положения солнца. Например, у голубиного гороха, Cajanus cajan, листочки, на которые падает солнце, ориентированы приблизительно параллельно солнечным лучам, что минимизирует площадь поверхности, подверженной интенсивному тропическому излучению. Но листья в тени ориентируются под прямыми углами к падающим лучам, тем самым улавливая максимальное количество света. Эти реакции зависят от направления и интенсивности света, падающего на специализированные узлы в листьях (листовые подушечки). В течение дня листья и листочки непрерывно приспосабливают свое положение к движению солнца по небу. Ночью они принимают свое вертикальное «спящее» положение: листовые подушечки чувствительны к гравитации так же, как и к свету.
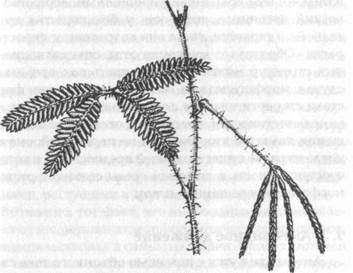
Рис. 23. Листья чувствительного растения, Mimosa pudica. Слева — не стимулированные; справа — стимулированные
У «чувствительного» растения, Mimosa pudica, листочки закрываются и листья смотрят вниз ночью, как и у многих других бобовых растений. Но эти движения быстро происходят также и в дневное время в ответ на механический стимул {рис. 23). Стимул вызывает распространение вниз по листу волны электрической деполяризации, подобной нервному импульсу; если стимул достаточно сильный, импульс распространяется на другие листья, которые также свертываются[188]. Подобно этому, у растения венерина мухоловка, Dionaea muscipula, механическая стимуляция чувствительных волосков на поверхности листа вызывает движение электрического импульса к разбухшим «шарнирным» клеткам, которые быстро теряют воду; в результате лист захлопывается как ловушка вокруг несчастных насекомых, которые затем перевариваются[189].
Эти движения листьев и листочков в ответ на действие света, силы тяжести и механическую стимуляцию возможны потому, что специализированные клетки способны терять воду, а затем вырастать снова; следовательно, они сохранили упрощенный морфогенетический потенциал, тогда как у большинства других тканей он теряется, когда они созревают и перестают расти. Обратимые движения этих специализированных структур можно рассматривать как предельные случаи морфогенеза, в которых изменения формы стали стереотипными и повторяющимися. Но их квазимеханистическая простота в эволюционном отношении является вторичной, а не первичной: она возникла из ранее существовавшей предпосылки, в которой чувствительность к стимулам среды связана с ростом и морфогенезом растения в целом.
Амебоидное движение
Амебы движутся с помощью объемного тока своей цитоплазмы в вырастающие выступы — псевдоподии. В норме они перемещаются по поверхности твердых объектов посредством непрерывного растягивания своих передних концов. Но если до псевдоподий дотронуться или если они соприкасаются с теплом или концентрированными растворами химических веществ, они перестают расти, вместо них вырастают другие, и тогда клетка меняет направление движения. Если новые псевдоподии снова встречают какие-либо потенциально опасные стимулы, они также останавливаются, и амеба уходит от них в другом направлении. Эта система «проб и ошибок» действует до тех пор, пока они не находят путь без препятствий или неблагоприятных стимулов[190].

Рис. 24. Метод, с помощью которого плавающая амеба переходит на твердую поверхность (согласно данным, приведенным в Jennings, 1906)
У свободно
|
|
|
