 |


Морфогенетические поля и моторные поля
|
|
|
|
Несмотря на то что поля, контролирующие изменения формы специализированных двигательных структур животных,— это фактически морфогенетические поля, они вызывают, скорее, движения, нежели изменения формы. По этой причине кажется более уместным называть их двигательными, или моторными, полями. (Слово «motor» употребляется здесь как прилагательное от существительного «motion» — движение.) Моторные поля, подобно морфогенетическим полям, зависят от морфического резонанса со стороны прошлых подобных систем, и их действие состоит в актуализации виртуальных форм. Канализованные пути к конечной форме или состоянию для моторных полей можно назвать хреодами, так же как и для морфогенетических полей.
Моторные поля, как и морфогенетические поля, имеют иерархическую организацию и, вообще говоря, связаны с развитием, выживанием и воспроизведением. В то время как у растений эти процессы почти полностью морфогенетичны, у животных они зависят также и от движения. Действительно, у большинства животных даже поддержание нормальных функций тела требует постоянного движения внутренних органов, таких как кишечник, сердце и дыхательная система.
В отличие от растений животные должны питаться другими живыми организмами, чтобы поддерживать свои формы. Поэтому важным общим для всех животных моторным полем является поле питания. Оно управляет вспомогательными полями, ответственными за нахождение, добычу и съедание растения или животного, которые служат пищей. Некоторые животные ведут сидячий образ жизни и заставляют еду двигаться к ним, например, с течениями воды; другие просто переходят с места на место, пока не находят растения, которые могут есть; третьи охотятся за другими животными; есть такие, которые делают ловушки, чтобы поймать жертву; некоторые являются паразитами; иные питаются падалью и так далее. Все эти способы питания зависят от иерархий специфических хреод.
|
|
|
Другой основной тип моторных полей связан с избеганием неблагоприятных условий. Amoeba и Рататесшт проявляют реакции наиболее простого типа: уход назад или в сторону от неблагоприятного стимула и движение в каком-либо ином направлении. Животные, ведущие сидячий образ жизни, такие как Stentor или Hydra, реагируют на слабый неблагоприятный стимул сокращением своего тела, но при более сильном стимуле они устремляются прочь и устраиваются где-нибудь в другом месте. В дополнение к общим реакциям избегания многие животные проявляют также особые виды поведения, которые позволяют им спасаться от хищников: например, они могут быстро убегать, или отстаивать свою позицию и как-либо пугать хищника, или застыть таким образом, что становятся менее заметными.
Конечной формой полных полей развития и выживания является взрослое животное, выросшее в оптимальных условиях. Когда бы ни достигалось это состояние, животному не нужно делать для этого что-либо особенное, но отклонения от такого состояния подвергают животное влиянию различных моторных полей, направленных к его восстановлению. В действительности такие отклонения происходят часто: непрерывный метаболизм у животного истощает его запасы пищи; изменения в окружающей среде ставят его в неблагоприятные условия; к нему неожиданно приближаются хищники. Эти и другие изменения обнаруживаются чувствительными структурами и вызывают характерные изменения нервной системы, которые затем становятся структурой зародыша для особого моторного поля.
Конечная форма полного поля воспроизведения есть создание жизнеспособного потомства. У одноклеточных организмов и у простых многоклеточных животных, таких как гидра, эта цель достигается с помощью морфогенетического процесса: организмы делятся надвое или выталкивают новые особи. Подобно этому, примитивные способы полового воспроизведения по существу морфогенетичны: многие низшие животные (например, морские ежи), а также низшие растения (например, морской сорняк Fucus) просто выбрасывают миллионы яйцеклеток и сперму в воду вокруг себя.
|
|
|
У более продвинутых животных сперма выделяется не наугад, но поблизости от яйцеклетки в результате специального брачного поведения. Таким образом полное поле воспроизведения начинает охватывать моторные поля поиска партнера, ухаживания, и спаривания. Организмы могут подпадать под влияние первого моторного поля в этом ряду вследствие внутренних физиологических изменений, сопровождающихся выделением гормонов, а также под действием обонятельных, визуальных или иных стимулов от потенциальных партнеров. Конечная точка первого поля создает зародыш для второго и так далее: за поиском партнера следует ухаживание, которое в случае успеха приводит к начальной точке хреоды спаривания. В простейших случаях конечная форма всей цепочки для мужского партнера есть извержение (семени), для женского — откладывание яиц. У многих водных организмов яйца просто откладываются в воду, но у земных животных откладывание яиц часто включает сложные и высокоспецифичные модели поведения: например, наездники (вид насекомого) откладывают свои яйца в гусеницы определенных видов, в которых развиваются личинки-паразиты, а осы-горшечники делают маленькие горшочки, куда они кладут парализованную жертву, затем откладывают на нее яйца и запечатывают «горшочки».
У некоторых живородящих видов потомство просто выталкивается наружу и оказывается покинутым сразу после рождения. Но когда о детенышах заботятся сразу после рождения или высиживания, начинает работать новая серия моторных полей, все еще находящихся под контролем полного поля воспроизведения родителей, но в то же время обслуживающих и поле развития и выживания детенышей. Следовательно, поведение животных приобретает социальное измерение. В простейших случаях такие сообщества носят временный характер и распадаются, когда потомство становится независимым; в других случаях сообщества сохраняются с соответствующим увеличением сложности поведения. Специальные моторные поля управляют различными видами общения между особями и разными задачами, которые выполняют отдельные индивидуумы.
|
|
|
В чрезвычайно сложных сообществах термитов, муравьев и общественных пчел и ос особи с подобным или идентичным генетическим строением выполняют совершенно различные задачи, а одно и то же насекомое даже может в разное время играть разные роли: например, молодая рабочая пчела может сначала чистить улей, затем, через несколько дней, «нянчить» выводящихся пчел, затем строить соты для меда, затем получать и укладывать пыльцу, затем охранять улей и, наконец, вылетать на поиски пищи[200]. Каждая из этих ролей должна направляться моторным полем высшего уровня, которое контролирует по очереди хреоды низших уровней, определяющие выполнение отдельных специальных задач. Изменения нервной системы насекомых должны ставить их под контроль одного или другого из этих полей высших уровней, способствуя их попаданию в морфический резонанс с предшествовавшими работниками, выполнявшими данную отдельную роль. Такие изменения до некоторой степени зависят от изменений в физиологии насекомого по мере его взросления, но на них сильно влияют также внешние стимулы: роли индивидов изменяются в ответ на повреждения улья или нарушения в пчелином сообществе; вся система регулируется.
Моторные поля высших уровней — питания, избегания, воспроизведения и т. д.— обычно управляют серией полей низших уровней, которые действуют последовательно, так что конечная форма одного обеспечивает зародышевую структуру для следующего. Моторные поля еще более низких уровней в этой иерархии часто действуют циклично, порождая повторяющиеся движения, такие как движения ног при ходьбе, крыльев в полете, челюстей при жевании. Самый низкий уровень занимают поля, во всех деталях контролирующие сокращение клеток в мускулах.
|
|
|
Моторные поля высших уровней охватывают не только органы чувств, нервную систему и мускулы, но также объекты вне животного. Рассмотрим, например, моторные поля питания. Полный процесс — поимка и заглатывание пищи — фактически представляет собой особый тип агрегативного морфогенеза (ср. раздел 4.1). Голодное животное является зародышевой структурой и вступает в морфический резонанс с предыдущими конечными формами этого моторного поля, а именно с полями подобных животных, существовавших в прошлом (включая его самого), в сытом состоянии. Если речь идет о хищнике, достижение этой конечной формы зависит от поимки и заглатывания добычи. Моторное поле поимки распространяется в пространстве вокруг животного и включает в него виртуальную форму добычи (рис. 11). Эта виртуальная форма актуализируется, когда сущность, достаточно близко ей соответствующая, приближается к хищнику: добыча узнается, и активизируется хреода поимки. Теоретически моторное поле может влиять на вероятностные события в любой или во всех системах, которые оно охватывает, включая органы чувств, мускулы и саму жертву. Но в большинстве случаев его влияние, вероятно, ограничено модификацией вероятностных событий в центральной нервной системе, что направляет движения животного к достижению конечной формы, в данном случае к поимке добычи.
Моторные поля и чувства
Посредством морфического резонанса животное попадает под влияние специфических моторных полей вследствие своей характерной структуры и внутренней организации колебательных процессов. Эти процессы изменяются в результате изменений, происходящих в теле животного, и влияний извне.
Если различные стимулы вызывали одинаковые изменения внутри животного, тогда начинают работать одни и те же моторные поля. По-видимому, это именно то, что происходит в одноклеточных организмах, которые проявляют одну и ту же реакцию избегания в ответ на разнообразные физические и химические стимулы: возможно, все они оказывают одинаковое воздействие на физико-химическое состояние клетки, например изменяя проницаемость клеточной мембраны для кальция или других ионов.
У простых многоклеточных животных с относительно слабой сенсорной специализацией диапазон реакций на стимулы ненамного больше, чем у одноклеточных. Например, гидра демонстрирует одинаковые реакции избегания на множество различных физических и химических стимулов и отвечает на объекты, такие как частицы пищи, только при механическом контакте. Однако, как и у некоторых одноклеточных организмов, ее реакция на твердые объекты изменяется под влиянием химических стимулов. Это можно показать на простом эксперименте: если к щупальцам голодной гидры подаются маленькие кусочки фильтровальной бумаги, реакции не наблюдается; но если они предварительно смочены в мясном соусе, щупальца несут их в рот, и затем они проглатываются[201].
|
|
|
Напротив, животные, имеющие глаза, которые формируют образы, могут чувствовать объекты, еще находящиеся на некотором расстоянии от них; следовательно, моторные поля здесь распространяются дальше в окружающую среду; диапазон и разнообразие поведения животных значительно возрастают. Подобным же образом чувство слуха позволяет обнаруживать удаленные объекты и поэтому позволяет расширить пространственную протяженность моторных полей даже до тех областей, где объект не может быть виден. У некоторых животных, особенно у летучих мышей, это чувство заменило зрение как основу протяженных моторных полей. А у некоторых видов, живущих в воде, таких как электрические рыбы (виды Mormyrid и Gimnotid), специализированные рецепторы обнаруживают изменения электрического поля вокруг них с помощью импульсов, испускаемых их электрическими органами; это позволяет им определять местонахождение добычи и других объектов в загрязненных тропических реках, где они обитают.
Когда животные движутся, чувственные стимулы, возникающие как внутри их тел, так и под влиянием среды, изменяются в результате их собственных движений. Эта непрерывная обратная связь играет существенную роль в координации движений их моторными полями.
Подобно морфогенетическим полям, моторные поля являются вероятностными структурами, которые с помощью морфического резонанса связываются с физическими системами через их трехмерные колебательные структуры. Поэтому фундаментальное значение имеет тот факт, что все колебательные вводы переводятся в пространственно-временные структуры, в которых осуществляется деятельность нервной системы. В чувстве осязания стимулы действуют на определенные участки тела, которые отмечены («картарованы») в мозгу в результате действия особых нервных путей; в зрении образы, попадающие на сетчатку, вызывают распределенные в пространстве изменения в оптических нервах и зрительной коре; и хотя обонятельные, вкусовые и слуховые стимулы не носят непосредственно пространственный характер, нервы, которые они возбуждают через соответствующие органы чувств, находятся в определенных местах и импульсы, проходящие по этим нервам в центральную нервную систему, создают характерные объемные распределения возбуждения.
Таким образом, отдельные стимулы и их комбинации производят характерные пространственно-временные эффекты. Эти динамические картины активности приводят нервную систему в морфический резонанс с подобными прошлыми нервными системами в аналогичных состояниях, и, следовательно, она попадает под влияние особых моторных полей.
Регуляция и регенерация
Подобно морфогенетическим полям, моторные поля направляют системы, находящиеся под их влиянием, к характерным конечным формам. Обычно они достигают этого, стимулируя серию движений в определенной последовательности. Промежуточные стадии стабилизируются с помощью морфического резонанса, другими словами, они являются хреодами. Но хреоды представляют собой просто наиболее вероятные пути к конечным формам. Если нормальный путь заблокирован или если система отклонилась от него по какой-либо причине, та же самая конечная форма может быть достигнута другим путем: система регулируется (раздел 4.1). Многие, но не все морфогенетические системы способны к регуляции; таковы и двигательные системы.
Регуляция происходит под действием моторных полей на всех иерархических уровнях: например, если несколько мускулов или нервов в ноге у собаки повреждены, организация сокращений в других мускулах регулируется таким образом, что конечность функционирует нормально. Если нога ампутирована, движения оставшихся ног изменяются так, что собака все же может ходить, хотя и хромает. Если повреждены части коры ее головного мозга, через некоторое время он восстанавливается более или менее полно. Если ее ослепить, ее способность двигаться постепенно улучшается, по мере того как она начинает более полагаться на оставшиеся чувства. А если прегражден обычный путь к дому, ее еде или щенкам, она изменяет привычную последовательность движений, пока не находит новый путь для достижения цели.
Поведенческий эквивалент регенерации встречается тогда, когда конечная форма хреоды была актуализирована, но затем разрушена: представьте, например, кота, который поймал мышь, что является конечной точкой хреоды поимки добычи. Если мышь ускользает из его когтей, то движения кота направлены на то, чтобы снова ее поймать.
Из всех примеров поведенческой регенерации ее соответствие регенерации морфогенетической лучше всего обнаруживается в морфогенетическом поведении, связанном с созданием характерных структур. В некоторых случаях животные улучшают эти структуры после их повреждения. Например, было сделано наблюдение, что осы-горшечники иногда заполняют отверстия, сделанные экспериментатором в стенках их горшочков, с помощью действий, которые они никогда не совершают при нормальном построении горшочков[202]. А термиты чинят повреждения в своих галереях и гнездах кооперативными и координированными усилиями множества отдельных насекомых[203].
Эти и подобные действия иногда интерпретировались как свидетельства существования разума на том основании, что животные, действующие строго фиксированным образом, задаваемым инстинктом, не были бы способны столь гибко реагировать на необычные ситуации[204]. Но, следуя этой логике, можно было бы сказать, что регулирующиеся эмбрионы морских ежей и регенерирующие плоские черви также проявляют признаки разума. Однако такое расширение физиологической терминологии, скорее, внесло бы путаницу, нежели оказалось полезным. С точки зрения гипотезы формативной причинности такие подобия признаются, но интерпретируются иначе. Способность животных достигать поведенческих целей необычными путями, рассматриваемая с позиций морфогенетической регуляции и регенерации, не дает оснований вводить новые фундаментальные принципы. А когда у высших животных некоторые типы поведения более не следуют стандартным хреодам — когда поведенческая регуляция становится, так сказать, скорее, правилом, чем исключением,— эту подвижность можно рассматривать как расширение возможностей, заложенных в самой природе морфогенетических и моторных полей.
Глава 10
Инстинкт и обучение
Влияние прошлых действий
Подобно морфогенетическим полям, моторные поля задаются морфическим резонансом от прошлых подобных систем. Детали строения животного и организация колебательной активности его нервной системы обычно подобны таковым у него самого более, нежели у какого-либо другого животного. Таким образом, наиболее специфическим морфическим резонансом, действующим на данное животное, будет резонанс от его собственной формы в прошлом (ср. раздел 6.5). Следующий наиболее специфический резонанс будет от генетически подобных животных, которые жили в тех же условиях, и наименее специфический — от животных других пород, живущих в других условиях. В «долинной» модели хреоды {рис. 5] последний стабилизирует общую форму, тогда как более специфический резонанс определяет детали топологии дна этой долины.
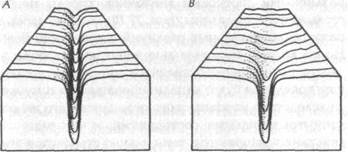
Рис. 27. Схематическое изображение глубоко канализированной хреоды (А) и хреоды, слабо канализированной на начальных стадиях (В)
«Контуры» долины хреод зависят от степени подобия между поведением родственных животных той же породы или вида. Если их модели движения варьируются слабо, морфический резонанс порождает глубокие и узкие хреоды, представленные долинами с крутыми краями {рис. 27 А). Такие хреоды оказывают сильное канализирующее действие на поведение последующих индивидуумов, которые по этой причине будут стремиться вести себя весьма сходным образом. Стереотипные модели поведения, обусловленные такими хреодами, на низших уровнях проявляются как рефлексы, а на высших уровнях — как инстинкты.
С другой стороны, если подобные друг другу животные достигают конечных форм своих моторных полей различными способами движения, хреоды не будут столь четко определены {рис. 27 В); поэтому здесь диапазон индивидуальных различий в поведении больше. Но после того как отдельное животное достигло цели своего поведения собственным путем, его последующее поведение будет канализироваться по тому же пути вследствие морфического резонанса с его же собственными прошлыми состояниями; и чем чаще такие действия повторяются, тем сильнее становится эта канализация. Такие характерные индивидуальные хреоды проявляют себя как привычки.
Таким образом, с точки зрения гипотезы формативной причинности между инстинктами и привычками разница лишь в степени: и те и другие зависят от морфического резонанса, первые — с бесчисленными предшествовавшими индивидуумами того же вида, а вторые — преимущественно с прошлыми состояниями того же индивидуума.
Этим мы не хотим сказать, что рефлексы и инстинкты не зависят от весьма специфически организованного морфогенеза нервной системы. Очевидно, что такая зависимость существует. Мы не хотим также сказать, что в процессах обучения не происходят физические или химические изменения в нервной системе, которые облегчают повторение данного вида движения. Возможно, в простой нервной системе, осуществляющей стереотипные функции, потенциал для таких изменений может быть уже «встроен» в систему «проводов» таким образом, что обучение происходит квазимеханически. Например, было обнаружено, что у улитки Aplysia строение нервной системы почти одинаково у разных особей, вплоть до мельчайших деталей расположения возбуждающих и ингибирующих синапсов на отдельных клетках. Очень простые виды обучения встречаются в связи с рефлекторным втягиванием жабер во впадину под покровной пластинкой, а именно привыкание к безвредным и приобретение чувствительности к опасным стимулам; при этих процессах функционирование отдельных возбуждающих и ингибирующих синапсов, действующих на индивидуальные клетки, определенным образом изменяется[205]. Конечно, простое описание этих процессов само по себе не выявляет причин таких изменений; в настоящее время можно лишь строить предположения на этот счет. Одно из них состоит в том, что эти модификации химического происхождения и связаны, вероятно, с изменениями в фосфорилировании белков[206]. Но как возникла эта тонкая специализация структуры и функций нервов и синапсов? Проблема переносится в область морфогенеза.
Нервные системы высших животных значительно больше варьируются от индивидуума к индивидууму, чем у беспозвоночных типа Aplysia, и они гораздо сложнее. Очень мало известно о том, каким образом сохраняются заученные способы поведения[207], но накоплено уже достаточно сведений, чтобы утверждать, что здесь не может быть простого объяснения на уровне специфически локализованных физических и химических «следов» в нервной ткани.
Многочисленные исследования показали, что у млекопитающих привычки, образовавшиеся при обучении, часто сохраняются после значительного повреждения коры и участков подкорки головного мозга. Более того, когда происходит потеря памяти, она не является тесно связанной с местонахождением таких повреждений, но зависит, скорее, от общего количества поврежденной ткани. К. С. Лэшли суммировал результаты сотен экспериментов следующим образом:
«Невозможно продемонстрировать изолированный участок как след памяти где-либо в нервной системе. Ограниченные области могут быть существенными для обучения или сохранения определенной активности, но в таких областях их части в функциональном отношении эквивалентны» [208].
Подобный же феномен был продемонстрирован на беспозвоночном — осьминоге: наблюдения сохранения приобретенных привычек после разрушения различных частей вертикальной доли мозга привели к кажущемуся парадоксальным выводу, что «память находится везде одновременно и нигде в частности»[209].
С механистической точки зрения эти результаты сильно озадачивают. В попытках найти им объяснение было высказано предположение, что «следы» памяти каким-то образом распределены в мозгу способом, аналогичным тому, который используется при сохранении информации в виде интерференционных картин в голограмме[210]. Но это пока не более чем неопределенное соображение.
Гипотеза формативной причинности дает альтернативное объяснение, в котором сохранение приобретенных привычек, несмотря на повреждение мозга, выглядит гораздо менее загадочно: привычки зависят от моторных полей, которые вообще не сохраняются в мозгу, но приходят непосредственно от его прошлого состояния путем морфического резонанса.
Некоторые приложения гипотезы формативной причинности применительно к проблемам инстинкта и обучения рассматриваются в следующих разделах, а в главе 11 предлагаются способы, с помощью которых можно экспериментально отличить предсказания, вытекающие из этой гипотезы, от предсказаний механистической теории.
Инстинкт
У всех животных некие модели двигательной активности являются, скорее, врожденными, нежели приобретенными в результате обучения. Наиболее фундаментальный характер имеют движения внутренних органов, таких как сердце и кишечник, но многие из способов движения конечностей, крыльев и других двигательных структур также являются врожденными. Это наиболее очевидно, когда животные оказываются способными совершать правильные, полезные для них движения почти сразу же после того, как они родились или вылупились из яйца.
Не всегда легко провести различие между врожденным и «обученным» поведением. Вообще говоря, характерное поведение, которое вырабатывается у молодых животных, воспитанных в изоляции, обычно может рассматриваться как врожденное; с другой стороны, поведение, которое появляется только при контакте с другими особями того же вида, также может быть врожденным, но требуются стимулы от других животных, чтобы оно проявилось.
Исследования инстинктивного поведения многих видов животных привели к нескольким общим выводам, которые составляют классические принципы этологии[211]. Их можно суммировать следующим образом:
(1) Инстинкты организованы в виде иерархии систем или центров, наложенных друг на друга. Каждый уровень активируется главным образом системой вышележащего уровня. Самый высокий центр каждого из основных инстинктов может испытывать влияние многих факторов, таких как гормоны, чувственные стимулы изнутри животного и стимулы из окружаю щей среды.
(2) Поведение, которое происходит под влиянием основных инстинктов, часто состоит из цепей более или менее стереотипных моделей поведения, называемых фиксированными моделями действия. Когда такая фиксированная модель создает конечную точку главной или малой цепи инстинктивного поведения, ее называют завершающим актом (consummately act). Поведение в ранней части инстинктивной цепи поведения, напри мер поиски пищи, может быть более гибким и называется обычно поведением инстинктивной потребности (appetitive behavior).
(3) Для активации, или «освобождения» (release), каждой системе требуется специфический стимул. Этот стимул, или «освободитель» (релизер), может при ходить изнутри тела животного или из окружающей среды. В последнем случае его часто называют знаковым стимулом. Предполагается, что данный релизер, или знаковый стимул, воздействует на специфический нейросенсорный механизм, называемый врожденным релизорным механизмом, который дает выход реакции.
Эти положения очень хорошо согласуются с идеей моторных полей, развивавшейся в предыдущей главе. Фиксированные модели действий находят объяснение на языке хреод, а врожденные освобождающие механизмы можно представить как структуры зародышей соответствующих моторных полей.
Знаковые стимулы
Инстинктивные отклики животных на знаковые стимулы показывают, что они как-то выделяют специфические и повторяющиеся особенности в своем окружении.
«Животное "слепо" отвечает только на одну часть окружающей его ситуации и пренебрегает другими ее частями, хотя его органы чувств прекрасно могут их воспринимать....Эти эффективные стимулы можно легко обнаружить, проверяя реакцию на разные ситуации, отличающиеся одним или другим из возможных стимулов. Более того, когда орган чувств участвует в "освобождении" реакции, лишь часть стимулов, которые он может получить, действительно являются эффективными. Как правило, инстинктивная реакция отвечает лишь на очень немногие стимулы, а большая часть окружения оказывает незначительное или не оказывает никакого влияния, даже если животное имеет чувствительные структуры для получения многочисленных деталей» (Н. Тимберген) [212].
Эти принципы иллюстрируются следующими примерами[213]. Агрессивная реакция самца рыбы колюшки в отношении других самцов в сезон размножения высвобождается главным образом знаковым стимулом красного брюшка: модели очень грубой формы, но с красными брюшками атакуются гораздо чаще, чем модели правильной формы, но без красной окраски.
Подобные же результаты были получены в экспериментах на красногрудой малиновке: охраняющий территорию самец ведет себя угрожающе по отношению к весьма приблизительным моделям с красными грудками или даже просто к пучку красных перьев, но гораздо слабее реагирует на точные модели птиц без красных грудок.
Молодые утки и гуси инстинктивно реагируют на приближение хищных птиц, причем эта реакция зависит от формы птицы в полете. Опыты с моделями из картона показали, что наиболее важным признаком является короткая шея, характерная для ястребов и других хищных птиц, тогда как размер и форма крыльев и хвоста имеют сравнительно мало значения.
У некоторых мотыльков сексуальный запах, или феромон, который обычно производят самки, заставляет самцов делать попытки к спариванию с любым объектом, обладающим таким запахом.
У саранчи вида Ephippiger ephippiger самцы привлекают самок, желающих спариваться, своей песней. Самки привлекаются к поющим самцам со значительного расстояния, но молчащих самцов они игнорируют, даже если те находятся поблизости. Самцы, которых заставляют молчать, склеивая их крылышки, неспособны привлекать самок.
Курицы приходят на помощь цыплятам в ответ на их крик о бедствии, но не тогда, когда они просто видят цыплят в беде, например за звуконепроницаемым стеклянным барьером.
Согласно гипотезе формативной причинности, распознавание этих знаковых стимулов должно зависеть от морфического резонанса от прошлых подобных животных, подвергнутых тем же стимулам. Благодаря процессу автоматического усреднения этот резонанс будет усиливать только общие черты пространственно-временных моделей активности, вызываемых этими стимулами в нервной системе. Результат состоит в том, что из окружения выделяются лишь некоторые специфические стимулы, тогда как другие игнорируются. Рассмотрим, например, стимулы, действующие на кур, чьи цыплята попали в беду. Вообразим набор фотографий цыплят в беде во многих различных случаях. Фотографии, сделанные ночью, не отобразят ничего; снятые в дневное время покажут цыплят разных размеров и форм, видимых спереди, сзади, сбоку или сверху; более того, они могут быть вблизи других объектов всех форм и размеров или даже спрятаны за ними. Далее, если негативы всех этих фотографий наложить друг на друга, чтобы получить составное изображение, в нем не будут усилены какие-либо черты, результатом будет просто расплывшееся пятно. Теперь вообразите, наоборот, магнитофонные записи, сделанные одновременно с фотографиями. На всех будут крики о бедствии, и если эти звуки накладываются друг на друга, они усиливают друг друга, давая в результате автоматически усредненный крик о бедствии. Это наложение фотографий и магнитофонных записей аналогично эффектам морфического резонанса от нервных систем предыдущих кур с последующей курицей, испытывающей стимул в виде крика цыпленка в беде: зрительные стимулы не возбуждают специфического резонанса и не вызывают инстинктивной реакции, каким бы несчастным ни выглядел цыпленок для наблюдающего человека, в то время как на слуховые стимулы реакция есть.
Этот пример иллюстрирует то, что является, по-видимому, общим принципом: формы очень часто неэффективны в качестве знакового стимула. Возможная причина в том, что они сильно варьируются, поскольку зависят от угла, под которым рассматривается объект. Напротив, цвета гораздо менее зависимы от точки зрения, а звуки и запахи вряд ли вообще зависят от нее.
Существенно, что цвета, звуки и запахи играют важную роль как «освободители» инстинктивных реакций; а в тех случаях, когда оказывается эффективной форма, имеется некоторое постоянство точки наблюдения. Например, птенцы на земле видят хищников, летающих над ними, как силуэты и действительно реагируют на такие формы. А когда формы, или модели, поведения служат сексуальными знаковыми стимулами, они делают это в сценах ухаживания, или в «представлениях», в которых животные принимают различные позы относительно своих потенциальных партнеров. То же справедливо для демонстрации покорности или агрессивных намерений.
Обучение
Можно сказать, что происходит обучение, когда имеется какое-либо относительно постоянное адаптивное изменение в поведении в результате прошлого опыта. Здесь можно выделить четыре общие категории[214]:
(1) Наиболее универсальным типом, который обнаруживается даже у одноклеточных организмов[215], является привыкание, которое можно определить как ослабление реакции в результате повторения стимула, не сопровождающегося каким-либо подкреплением его значимости. Известный пример такого рода — исчезновение реакций тревоги или избегания на новые стимулы, которые оказываются безвредными: животные к ним привыкают.
Этот феномен предполагает существование своего рода памяти, позволяющей узнавать стимулы, когда они повторяются. По гипотезе формативной причинности это узнавание обусловлено главным образом морфическим резонансом организма с его собственными прошлыми состояниями, включая те, которые были вызваны новыми сенсорными стимулами. Этот резонанс служит для поддержания и фактически для определения идентичности организма с самим собой в прошлом (раздел 6.5). Повторяющиеся стимулы от окружения, отклики на которые не подкрепляются, станут действующей частью собственного «фона» организма. Наоборот, любые новые особенности окружения будут выделяться, поскольку они не распознаются как «свои»: обычно реакцией животного будет тревога или избегание именно потому, что стимулы ему незнакомы.
В случае некоторых стереотипных ответов, — таких как рефлекс отдергивания рожек у улитки Aplysia, привыкание может происходить квазимеханистическим образом на основе предсуществующих структурной и биохимической специализаций в нервной системе (раздел 10.1). Но если так, эта специализация является вторичной и возникает, вероятно, в результате ситуации, в которой привыкание более непосредственно зависит от морфического резонанса.
(2) У всех животных врожденные модели двигательной активности выявляются по мере того, как индивид взрослеет. В то время как одни прекрасно действуют в первый же раз, когда они выполняются, другие совершенствуются со временем. Например, первые попытки птенца взлететь или детеныша млекопитающего — ходить могут быть успешными лишь отчасти, но они улучшаются после повторных усилий. Не все такие улучшения обусловлены практикой: в некоторых случаях это просто результат созревания и происходит в такой же степени с течением времени у животных, которые были иммобилизованы[216]. Тем не менее многие виды двигательных приемов улучшаются таким способом, который нельзя приписать созреванию.
С точки зрения гипотезы формативной причинности этот тип обучения можно интерпретировать как регуляцию поведения. Морфический резонанс от бесчисленных прошлых особей данного вида автоматически дает усредненную хреоду, которая руководит первыми попытками животного осуществить определенную врожденную модель движения. Эта стандартная хреода может дать лишь приблизительно удовлетворительные результаты, например из-за отклонений от нормы органов чувств у животного, или его нервной системы, или двигательных структур. По мере того как движения совершаются, регуляция самопроизвольно вызывает к жизни «тонкие подстройки» к общей хреоде, а также к хреодам низшего уровня, которые она контролирует. Эти «подстроенные» хреоды будут стабилизироваться путем морфического резонанса с прошлыми состояниями самого животного, по мере того как будет повторяться эта модель поведения.
(3) Животные могут начать отвечать на какой-либо стимул реакцией, которая в норме вызывается другим стимулом. Такой тип обучения имеет место тогда, когда новый стимул действует одновременно или сразу же после начального стимула. Классическими примерами явля
|
|
|
