 |


Антителообразование. Рис. 63. Трансформация В-лимфоцитов в антителообразующие. клетки
|
|
|
|
Антителообразование
Образование антител как иммунная реакция на антигены происходит в лимфоидной ткани периферических органов иммунитета, главным образом, в лимфатических узлах и белой пульпе селезенки. Продуцентами антител являются плазмоциты.
Общая иммунная реакция на антиген. Синтез антител начинается в тимуснезависимой зоне и в стенках лимфатических синусов, сопровождаясь общим нарастанием клеточности лимфоузлов, увеличением в них центров размножения с большим количеством митотически делящихся лимфоцитов, появлением вокруг них и в медуллярных тяжах плазматических клеток. В первые сутки после введения антигена резко снижается выход лимфоцитов из лимфатических узлов, а в последующие 3-4 дня, наоборот, значительно возрастает, что ведет к интенсивной миграции (расселению) стимулированных лимфоцитов через кровь во все лимфоидные ткани.
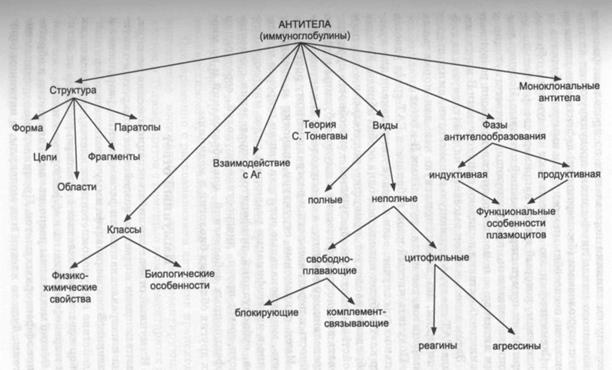
Фазы антителообразования. В динамике образования антител различают две фазы - индуктивную (латентную) и продуктивную, или репродуктивную.
Индуктивной фазой называют отрезок времени между введением антигена и появлением следов иммуноглобулинов или первых плазмоцитов. В этой фазе происходит распознавание антигена. Он фагоцитируется макрофагами (пиноцитируется) или связывается ими. Если вслед за этим антиген полностью разрушается, то антитела не вырабатываются. Антителогенез происходит лишь при частичной деградации антигена. В таком случае в лимфоидной ткани, где происходит связывание антигена, отмечаются массовая гибель и, параллельно, пролиферация клеток, появляется большое количество фагоцитов, увеличивается содержание гистамина, гепарина, серотонина и других веществ, усиливающих воспаление. На этом фоне в конце индуктивной фазы начинается взаимодействие (кооперация) между вспомогательными А-клетками, на мембране которых находится измененный антиген, Т- и В-лимфоцитами, в результате чего молодые В-лимфоциты, или предшественники плазмоцитов, получают сигнал к пролиферации и дифференциации (рис. 63).
|
|
|
 Индуктивная фаза очень лабильна и длится примерно 20 ч. Начавшийся процесс антителообразования в ней можно приостановить воздействием неблагоприятных для организма факторов. Легче всего это удается с помощью радиации, вследствие чего индуктивную фазу антителогенеза называют радиочувствительной.
Индуктивная фаза очень лабильна и длится примерно 20 ч. Начавшийся процесс антителообразования в ней можно приостановить воздействием неблагоприятных для организма факторов. Легче всего это удается с помощью радиации, вследствие чего индуктивную фазу антителогенеза называют радиочувствительной.
Рис. 63. Трансформация В-лимфоцитов в антителообразующие
клетки
В продуктивной фазе происходит интенсивный синтез антител. Приостановить его не удается даже радиооблучением, и поэтому продуктивную фазу называют радиорезистентной. В этой фазе иммунного ответа усиливается кооперация иммунокомпетентных клеток, и сильно возрастает деление В-лимфоцитов. На смену молодым плаз-мобластам (иммунобластам) в лимфоидной ткани приходят юные плазмоциты, а затем - и зрелые.
При первичном иммунном ответе максимальное количество плазмоцитов в селезенке и лимфатических узлах появляется на 7-е сут. Это совпадает с высоким титром антител в крови. Повторное введение чужеродного антигена сопровождается необычно интенсивным антителообразо-ванием - высокий титр антител возникает уже на 3-4-е сут, что связано с наличием в организме первично иммунизированных особей клеток иммунологической памяти, не закончивших полный цикл трансформации (см. «Иммунный ответ»).
Продуктивная фаза антителообразования сравнительно непродолжительна. Применительно к одному клону она продолжается около 10 сут, но по отношению ко многим из них - может удлиняться до нескольких недель. Однако через 2-3 мес. титр антител в крови переболевших инфекционными заболеваниями резко снижается.
|
|
|
Функциональные особенности плазмоцитов. Касаясь функциональных особенностей плазмоцитов, следует подчеркнуть, что они, в известном смысле, могут рассматриваться как своеобразные одноклеточные белковые железы. Плазмоциты секретируют иммуноглобулины по голокриновому или мерокриновому типу. Как правило, плазмоциты образуют антитела одной иммунологической специфичности, например Н- или О- антитела к соответствующим жгутиковым и соматическим антигенам бактерий. Более того, при наличии в молекуле антигена двух разных детерминант плазмоцит вырабатывает антитела против одной из них. Только 0, 01 % плазмоцитов продуцирует оба антитела. Большинство плазмоцитов синтезирует лишь один класс тяжелых и один тип легких цепей одновременно. Первичный иммунологический ответ обычно начинается с синтеза макроглобулинов М с константой седиментации 19S. При вторичном ответе на антиген образуются микроглобулины G с константой 7S. Большая часть плазматических клеток образует антитела с константой седиментации либо 7S, либо 19S.
Развитие клона плазмоцитов, начиная от плазмобласта до зрелой формы, занимает 5-6 сут. Жизненный цикл плазмоцитов, продуцирующих тот или иной вид антител, не превышает 48 ч. При этом вырабатывающиеся клоном плазмоцитов антитела представляют собой широкий спектр иммуноглобулинов, отличающихся как по специфичности по отношению к различным детерминантам антигена, так и по авидности.
Моноклональные антитела. Методику получения высокоспецифических антител с большей степенью авидности к определенной детерминантной группе антигена разработали Ц. Милыптейн и Г. Келер, создав бессмертные клоны В-клеток, которые называют гибридомами. Гибридомы получают путем слияния нормальных по продолжительности жизненного цикла лимфоцитов, продуцирующих антитела, с опухолевыми (бессмертными) дефектными линиями В-клеток, неспособными к секреции иммуноглобулинов.
Схематически это осуществляют следующим образом. Антигеном, который имеет, например, две или больше детерминант, иммунизируют мышей или крыс. Получив лимфоциты селезенки, продуцирующие соответствующие к ним антитела, с помощью полиэтиленгликоля разрушают их клеточные стенки и сливают с опухолевыми В-клетками.
|
|
|
Затем взвесь клеток высевают на элективную для гибрида ГАТ-среду, содержащую гипоксантин, аминоптерин и тимидин, но неблагоприятную как для нормальных лимфоцитов, так и для опухолевых В-клеток.
Получить чистый клон гибридом в ГАТ-среде удается только в том случае, если в лунке микропанели для культур клеток будет находиться одна особь гибрида. Каждая гибридома наследует присущие родительским клеткам свойства: от антителообразующей - способность продуцировать высокоспецифические иммуноглобулины к строго определенному эпитопу, а от опухолевой В-клетки — способность к перевиваемости и неограниченному росту.
Имея одинаковые структуру, идиотип и специфичность, моноклональные антитела нашли широкое применение в идентификации различных видов микроорганизмов, субпопуляций лимфоцитов, определении совместимости групп крови, диагностике опухолей, серотерапии инфекционных заболеваний, новообразований и во многих других областях биологии и медицины.
Иммунный ответ
Иммунный ответ - это антигенспецифическая реакция, развивающаяся в ответ на воздействие конкретного антигена и направленная на его распознавание, разрушение и удаление. В основе развития иммунного ответа лежит взаимодействие иммунокомпетентных клеток, определяющих выбор клеток, включаемых в ответную реакцию на антиген и трансформацию их в эффекторные клетки.
Общие механизмы распознавания антигенов и образование эффекторных клеток. Проникающие в организм антигены, будь то клетки или молекулы, в нативном (природном) виде не распознаются как генетически чужеродные метки (исключением являются В-лимфоциты). Иммунная реакция их распознавания начинается с фагоцитоза антигена макрофагами крови и тканей или с его связывания с дендритными клетками стромы лимфоидных органов. В макрофагах они могут полностью разрушаться, но чаще расщепляются до промежуточных продуктов. В частности, большинство антигенов в течение часа в лизосомах фагоцитов подвергаются ограниченной денатурации и про-1еолизу или, как чаще говорят, процессингу. Оставшиеся в них пептиды (9-10 - 12-25 остатков аминокислот), называемые номинальным антигеном, комплексируются с репрессированными на внешней мембране макрофагов молекулами МНС класса II и в таком состоянии представляются для распознавания. В дендритных клетках антиген тоже процессируется, но может оставаться, как на па-нхиматозных клетках, в неизмененном виде. В случае процессинга переструктурированный в них антиген комплексируется с молекулами МНС класса II и класса I. Макрофаги, дендритные и некоторые другие вспомогательные клетки, несущие на внешней мембране комплек-молекул МНС с включенными в них пептидами, предназначенные для распознавания антигена, называются антигенпрезентирующими (presentatio - предъявление), мембранные комплексы антигенпрезентирующих клеток познают Т-лимфоциты. Таким образом, Т-лимфоциты познают не антиген как таковой, а его фрагменты, изменяющие структуру антигенов главного комплекса гистосовместимости, т. е. «измененное свое».
|
|
|
Одновременно с распознаванием антигена происходит контактное взаимодействие антигенпрезентирующих клеток и Т-лимфоцитов с помощью комплементарных поверхностных молекул, что обеспечивает их взаимную адгезию и секрецию антигенпрезентирующими клетками цитокинов, которые вызывают сначала активацию Т-лимфоцитов, а затем - пролиферацию и их дифференциацию в эффекторные клетки. К
Кооперация иммунокомпетентных клеток в гуморальном иммунном ответе. Иммунный ответ в виде антителообразования происходит при распознавании В-клеткой гена, который индуцирует ее пролиферацию и дифференциацию в плазмоциты. Прямое воздействие на В-клетку без участия Т-клеток могут оказывать только тимуснезависимые антигены. По механизму стимулирующего действия их разделяют на два типа: тип 1, к которому относятся бактериальные липополисахариды, и тип 2, включающий полисахариды стрептококков, полимеры D-аминокислот, поливинилпирролидон, полимеризованный флагеллин, не подвергающиеся деградации в лизосомах макрофагов. Тимуснезависимые антигены типа 1 стимулируют деление В-клеток неспецифически, воздействуя как митогены. Антигены типа 2 с многократно повторяющимися детерминантами реагируют с иммуноглобулино-выми рецепторами В-клеток. При этом в специфическом механизме активации В-клеток, кроме них, участвуют макрофаги, выделяя цитокины, стимулирующие их деление и трансформацию в плазмоциты.
|
|
|
Гуморальный иммунный ответ на тимусзависимые антигены начинается со связывания антигена рецептором В-лимфоцитов. Образующийся при этом на мембране В-лимфоцита комплекс IgM+Ar погружается внутрь клетки, где после процессинга и связывания с цитоплазматическими молекулами МНС класса II презентируется на его поверхности для распознавания Т-хелпером. Таким образом, в молекуле тимусзависимого антигена существует два набора эпитопов. Один из них распознается антигенспецифически-ми рецепторами BCR В-лимфоцитов, а второй, формируемый внутри В-клетки, распознается СБ4+-Т-хелперами. Эпитоп первого набора, распознаваемый В-клеткой, может иметь различную химическую структуру, например поли-сахаридную или белковую, а эпитоп второго набора, распознаваемый Т-хелперами, - только белковую. Естественно, что по отношению к эпитопам первого набора В-клетка является распознающей, а к эпитопам второго набора - презентирующей. В результате распознавания Т-хелпером эпитопов второго набора с помощью TCR-рецептора при участии корецептора CD4 и костимулирующих молекул CD28 и CD40L, взаимодействующих, как это показано на рис. 64, с соответствующими им молекулами CD80 и 40, Т-хелперы (ТхО) дифференцируются в Тх2-хелперы, выделяющие ИЛ-4, под влиянием которого В-клетка дифференцируется в плазмоцит, секретирующий антитела.
 Кооперация иммунокомпетентных клеток в цитотоксических реакциях. Цитотоксической называют иммунную реакцию, приводящую к формированию цитотоксических лимфоцитов (ЦТЛ), способных разрушать инфицированные или опухолевые (мутантные) клетки с экспрессируемым опознавательным антигеном на их поверхности. В качестве антигенпрезентирующих клеток, как это представлено на рис. 65, в ней участвуют дендритные клетки или макрофаги, презентирующие антигенный пептид в комплексе с молекулами МНС класса II Т-хелперам (С04+-клеткам), а в комплексе с МНС класса I - пре-Цитотоксическим лимфоцитам (СБ8+-клеткам). Для активации тех и других клеток обязательна костимуляция, обусловленная взаимодействием молекул CD80 антиген-презентирующих клеток с корецептором CD28 Т-лимфоцитов. Распознавшие презентированный антигенный пептид СБ4+-клетки продуцируют интерферон γ, активирующий антигенпрезентирующие клетки, а распознавшие антигенный пептид СБ8+-клетки подвергаются бласттрансформации, экспрессируя молекулы-рецепторы для ИЛ-2, который крайне необходим им для превращения в ЦТЛ и последующего размножения. Источником ИЛ-2 для ЦТЛ на первом этапе служат активированные вместе с ними ТхО-хелперы. Допускается, что на более поздних этапах ИЛ-2 начинают сами вырабатывать СВ8+-клетки, стимулированные антигенпрезентирующими клетками, т. е. обеспечивают свое размножение по аутокринному механизму.
Кооперация иммунокомпетентных клеток в цитотоксических реакциях. Цитотоксической называют иммунную реакцию, приводящую к формированию цитотоксических лимфоцитов (ЦТЛ), способных разрушать инфицированные или опухолевые (мутантные) клетки с экспрессируемым опознавательным антигеном на их поверхности. В качестве антигенпрезентирующих клеток, как это представлено на рис. 65, в ней участвуют дендритные клетки или макрофаги, презентирующие антигенный пептид в комплексе с молекулами МНС класса II Т-хелперам (С04+-клеткам), а в комплексе с МНС класса I - пре-Цитотоксическим лимфоцитам (СБ8+-клеткам). Для активации тех и других клеток обязательна костимуляция, обусловленная взаимодействием молекул CD80 антиген-презентирующих клеток с корецептором CD28 Т-лимфоцитов. Распознавшие презентированный антигенный пептид СБ4+-клетки продуцируют интерферон γ, активирующий антигенпрезентирующие клетки, а распознавшие антигенный пептид СБ8+-клетки подвергаются бласттрансформации, экспрессируя молекулы-рецепторы для ИЛ-2, который крайне необходим им для превращения в ЦТЛ и последующего размножения. Источником ИЛ-2 для ЦТЛ на первом этапе служат активированные вместе с ними ТхО-хелперы. Допускается, что на более поздних этапах ИЛ-2 начинают сами вырабатывать СВ8+-клетки, стимулированные антигенпрезентирующими клетками, т. е. обеспечивают свое размножение по аутокринному механизму.
 В отличие от Т-хелперов, нуждающихся для продолжительной пролиферации в контакте с АПК и в действии многих цитокинов, поддержание размножения активированных ЦТЛ обеспечивается одним ИЛ-2. Интересно, что, стимулируя размножение ЦТЛ, этот цитокин вместе с тем усиливает экспрессию к нему рецепторов и интенсифицирует дальнейшую его выработку. Завершившие деление ЦТЛ, ничем не отличающиеся по внешним признакам от преСБ8+-клеток, выселяются из паракортикальных зон лимфоузлов и, попав в циркуляцию, разносятся в различные органы и ткани, где могут разрушать инфицированные и перерождающиеся клетки.
В отличие от Т-хелперов, нуждающихся для продолжительной пролиферации в контакте с АПК и в действии многих цитокинов, поддержание размножения активированных ЦТЛ обеспечивается одним ИЛ-2. Интересно, что, стимулируя размножение ЦТЛ, этот цитокин вместе с тем усиливает экспрессию к нему рецепторов и интенсифицирует дальнейшую его выработку. Завершившие деление ЦТЛ, ничем не отличающиеся по внешним признакам от преСБ8+-клеток, выселяются из паракортикальных зон лимфоузлов и, попав в циркуляцию, разносятся в различные органы и ткани, где могут разрушать инфицированные и перерождающиеся клетки.
Главным результатом дифференциации ЦТЛ является реализация иммунного цитолиза. Отличается он от неиммунного, вызванного NK-киллерами, тем, что происходит через определенное время, необходимое для формирования активных Т-киллеров. Клетки-мишени распознаются такими киллерами по комплексам антигенных пептидов с молекулами МНС класса I, но без участия антигенпрезентирующих клеток. Механизм иммунного Т-клеточного Цитолиза почти таков же, как NK-зависимого, т. е. после установления тесной связи с клеткой-мишенью в зону их межклеточного контакта из гранул ЦТЛ поступают перфорин и гранзимы (фрагментины), которые после активации в присутствии ионов Са2+ перфорируют мембрану мишени и внутри клетки фрагментируют ДНК. В отличие от NK-зависимого, в клеточном цитолизе могут участвовать и другие цитотоксические вещества, например лимфотоксин и фактор некроза опухолей, а также выделяемый Т-хелперами и Txl-клетками интерферону, усиливающий экспрессию молекул гистосовместимости на поверхности клеток-мишеней, что способствует распознаванию чужеродного пептида.
Таким образом, в отличие от дифференциации лимфоцитов при созревании, в процессе которой формируются антигенраспознающие клетки, их дифференциация в иммунном ответе направлена на образование эффекторных клеток. Так, в гуморальном ответе происходит формирование крупных плазматических клеток с обильной цитоплазмой, и они переходят от синтеза мембранных иммуноглобулинов, имеющихся у В-лимфоцитов, к секреции растворимых lg, а при клеточном цитотоксическом ответе -цитотоксических лимфоцитов, синтезирующих вещества, разрушающие клетки-мишени, и цитокины, регулирующие активацию, рост и дифференциацию лимфоцитов и эффекторных клеток воспаления. При этом в гуморальном ответе в качестве антигенпрезентирующих клеток обычно выступают В-лимфоциты, а Т-хелперами служат Тх2-клетки, а в цитотоксическом - дендритные клетки и ТхО- или Txl-клетки.
Регуляторами иммунного ответа по-прежнему, можно считать, являются Т-супрессоры, которые в процессе образования иммуноглобулинов и в реакциях цитолиза блокируют выработку аутоантител и обеспечивают толерантность к собственным тканям и органам.
Клетки памяти. Параллельно с образованием плазмо-цитов и ЦТЛ формируются клетки памяти.
В-клетки памяти представляют собой особую линию дифференциации. Они образуются в зародышевых центрах вторичных фолликулов лимфатических узлов. Решающим моментом индукции В-клеток памяти является СБ40-опосредованное взаимодействие предшественников В-клеток с Т-хелперами. Их фенотип, если не считать экспрессии молекул CD45RO, практически не отличается от фенотипа обычных В-клеток. Рецепторы этих клеток обладают более высоким сродством к антигену, и они легко активируются, контактируя с Т-клеткой при представлении ей антигенного пептида и действии Т-клеточных цитокинов, что имеет значение для быстрого выхода В-клеток памяти в пролиферацию. Формирование В-клеток, ответственных за иммунологическую память, обеспечивающих быструю элиминацию антигенов и абсолютную устойчивость к реинфекции, отличается интересными закономерностями. В течение 10-20 дней после первичной иммунизации их количество неуклонно возрастает. Затем оно держится на постоянном уровне в течение нескольких месяцев, а иногда и лет. У клеток иммунологической памяти все направления синтеза иммуноглобулинов, за исключением одного, репрессированы, и для них антиген служит директивным включателем уже детерминированной пролиферации и дифференциации, которая заканчивается образованием плазмоцитов за 2-3 дня.
Менее изучены Т-клетки памяти. Так как они имеют все основные маркеры Т-лимфоцитов, включая рецептор-ный комплекс CD3-TCR, вспомогательные и адгезивные молекулы, то считается, что Т-клетки памяти относятся к их долгоживущим вариантам. Наиболее специфическим признаком Т-клеток памяти является экспрессия на их поверхности тех же молекул CD45RO и отсутствие CD45RA. В популяции Т-клеток памяти имеются как СБ4+-клетки, так и СВ8+-лимфоциты, но преобладают первые. Все они формируются в паракортикальной зоне лимфоузлов и параартериальных муфтах селезенки.

Цитокины
Общие свойства. Цитокинами (cytos - клетка, kineo - возбуждаю) называют вещества, активирующие и координирующие взаимодействие клеток. По структуре они являются гликопротеинами с молекулярной массой от 8 кД до 80 кД, а по эффекту действия - медиаторами. Вырабатываются при иммунном ответе, развитии воспаления и кроветворении, регулируя и стимулируя их. Обладают потенцирующим и аддитивным действием. Однако могут оказывать и противоположный эффект. Быстро синтезируясь (в течение 1-4-24 ч), они в короткие сроки расходуются или выводятся из кровотока через почки. При угасании иммунной реакции синтез цитокинов прекращается. Сродство к цитокинам определяется наличием на мембранах клеток специфических рецепторов, но многие из них отличаются плейотропизмом.
Продуценты цитокинов. Цитокины вырабатывают Т-лимфоциты, В-лимфоциты, NK-клетки, моноциты-макрофаги, дендритные и другие клетки соединительной ткани.
Классификация цитокинов. Цитокины лимфоцитов называют лимфокинами, моноцитов - монокинами, клеток стромы - гемопоэтинами {haima — кровь, poiesis - выработка). При этом лимфокины регулируют активацию, рост и дифференциацию лимфоцитов, монокины стимулируют неспецифические эффекторные клетки воспаления, а гемопоэтины - процессы кроветворения.
Независимо от того, какими клетками вырабатываются цитокины, они подразделяются на четыре группы: интерлейкины (inter - взаимно, leukos - белый), координирующие взаимодействие между лейкоцитами; интерфероны (ИФН), обладающие противовирусной активностью; факторы некроза опухолей (ФНО), разрушающие мутантные клетки (см. «Противоопухолевый иммунитет»); колоние-стимулирующие факторы (КСФ), вызывающие экстрамедуллярные (внемозговые) очаги кроветворения. Отдельно выделяют трансформирующий фактор роста Р (ТФРР ).
В норме без антигенной стимуляции иммунной системы в кровотоке обнаруживаются небольшие количества цитокинов, в основном цитокины стромальных клеток и моноцитов, стимулами для секреции которых могут, по-видимому, служить контакты с окружающими клетками, метаболиты и многие другие факторы. В отсутствие стимуляции лимфокины не выделяются, и покоящиеся лимфоциты обычно на них не реагируют. Активированные клетки начинают продуцировать цитокины в ближайшие часы. Так, монокины и гемопоэтины обнаруживаются спустя 1ч, интерлейкины - через 2 (ИЛ-2), 4 (ИЛ-4), 6 ч (ИЛ-10). Пик выработки моно- и гемопоэтинов приходится на 3-14-й ч, а большинства интерлейкинов - на 2-3 сут.
Биологическая характеристика цитокинов. В настоящее время описано около 20 интерлейкинов, два ФНО (α и β ), три ИНФ (α, β и γ ), три КСФ.
|
|
|
