 |


Таблица 5. Основные клетки-продуценты и их цитокины
|
|
|
|
Таблица 5. Основные клетки-продуценты и их цитокины
| Клетки- | Группы цитокинов | ||||
| продуценты | ил | ИФН | ФНО | КСФ | ТФР |
| ТхО | 2, 3, 4, 5, 6, 8, 9, 10, 13, 14 | γ | α, β | гм- | |
| Txl | 2, 3 | Y | α, β | гм- | |
| Тх2 | 3, 4, 5, 6, 9, 10, 13, 14 | α | гм- | ||
| цтл | 2, 3, 4, 5, 6, 9, 10, 13, 16 | У | α. β | гм- | |
| NK-клетки | 1, 8, 12 | У | β | гм- | |
| В-клетки | 1, 4, 10, 12 | а | α | ||
| Моноциты-макрофаги | 1, 3, 4, 5, 6, 7, 8, 10, 12, 15 | α, β | α | ГМ-, г- | β |
| Дендритные клетки | 1, 6, 12 | ||||
Как видно из табл. 5, наибольший спектр цитокинов синтезируют ТхО, но продуцируют они их в очень малом количестве. Более высокий уровень цитокинов секретиру-ют воспалительные Txl- и иммунные Тх2-лимфоциты. Цитотоксические лимфоциты образуют тоже большое количество цитокинов, но интенсивность их секреции намного ниже, чем у всех трех типов хелперов, моноцитов-макрофагов и клеток стромы.
Спектр действия интерлейкинов
Интерлейкины участвуют в различных типах межклеточных коммуникаций (связях). По спектру действия на клетки-мишени их можно подразделить на три группы: полифункциональные интерлейкины; участвующие в иммунном ответе; интерлейкины-хемоаттрактанты.
Полифунциональные интерлейкины. К группе полифункциональных интерлейкинов относятся шесть интер-лейкинов - ИЛ-1, ИЛ-3, ИЛ-6, ИЛ-7, ИЛ-9 и ИЛ-11. Все они имеют широкий спектр клеток-мишеней. Так, ИЛ-1 вызывает пролиферацию активированных В-клеток и их дифференциацию в плазмоциты; индуцирует воспалительную реакцию; повышает подвижность нейтрофилов; стимулирует фагоцитоз; способствует усилению синтеза белков острой фазы воспаления; усиливает продукцию других цитокинов; стимулирует ранние этапы эритропоэ-за; наконец, участвует в обмене сигналами между иммунной и нервно-эндокринной системами.
|
|
|
Столь же разнообразны клетки-мишени ИЛ-6 и ИЛ-11, которые по межсистемным взаимодействиям настолько сходны с ИЛ-1, что их трудно, а подчас и невозможно отличить. Клетками-мишенями трех остальных полифункциональных интерлейкинов являются в основном предшественники клеток крови (ИЛ-3 и ИЛ-9), ранние проВ- и преВ-клетки (ИЛ-7), тучные клетки (ИЛ-9).
Интерлейкины иммунного ответа. Среди интерлейкинов этой группы выделяют интерлейкины гуморального и клеточного ответов.
В гуморальном иммунном ответе участвуют ИЛ-4, ИЛ-5, ИЛ-13 и ИЛ-14. Основными из них являются ИЛ-4 и гомологичный ему ИЛ-13. Их мишенями являются В-лимфоциты. Действуя на них раньше других цитокинов, они могут стимулировать их размножение (при активации IgE имеют прямое отношение к развитию анафилаксии); являются основным фактором дифференциации ТхО-клеток в Тх2; повышают экспрессию молекул МНС класса II и антигенпрезентирующую активность вспомогательных клеток. ИЛ-5 и ИЛ-14, являясь ростовыми и дифференцировочными факторами В-лимфоцитов, вместе с тем стимулируют эозинофилы, потенцируя их гельмин-тоцидное действие (ИЛ-5), и индуцируют образование клеток памяти (ИЛ-14).
В клеточном цитотоксическом ответе принимают участие ИЛ-2 и близкие ему по структуре и функциям ИЛ-12 и ИЛ-15, которые необходимы как индукторы роста, пролиферации и дифференциации цитотоксических лимфоцитов (высокие их концентрации усиливают цитолити-ческий эффект NK-клеток), индуцируют образование на многих клетках специфических к ним рецепторов; воздействуя на моноциты, усиливают генерацию активных форм кислорода и пероксидов. ИЛ-12 примечателен тем, что является индуктором дифференциации Т-хелперов в направлении Txl и стимулятором выработки интерферона Y (см. «Иммунный ответ»).
|
|
|
Отдельного описания заслуживает ИЛ-10 как цитокин, участвующий в клеточно-гуморальном иммунном ответе. Так, совместно действуя с ИЛ-2 и ИЛ-7, он усиливает пролиферацию тимоцитов, а в сочетании с ИЛ-4 - ускоряет размножение В-клеток.
Интерлейкины-хемоаттрактанты. Истинными хемоат-трактантами среди цитокинов являются низкомолекулярный ИЛ-8 (7, 5 кД) и высокомолекулярный ИЛ-16 (80 кД). Первый из них повышает хемотаксис нейтрофилов, а второй - адгезивность Т-хелперов.
Факторы некроза опухолей
Имеется два фактора некроза опухолей: ФНОа и ФНОР (в прежней номенклатуре - лимфотоксины). ФНОа продуцируется в основном моноцитами-макрофагами, а ФНОР -Т-лимфоцитами, стимулированными различными антигенами и митогенами. Тот и другой факторы представляют собой гликопротеиды с молекулярной массой 17, 4 кД (ФНОа) - 20-25 кД (ФНОР). Действуют они на один и тот же рецептор, вызывая одинаковые эффекты. В частности, разрушают опухолевые и инфицированные вирусами и паразитами клетки с развитием в тканях геморрагического некроза и общей кахексии организма, вследствие чего еще недавно ФНО именовали кахексинами. Правда, степень их активности неодинакова. Более выражена она у ФНОа, что, может быть, связано со скоростью его образования. Так, в максимальном количестве он накапливается уже через 2-3 ч, а ФНОР - только на 2-3-и сут. По спектру клеток-мишеней и биологическим эффектам ФНО близки ИЛ-1 и ИЛ-6. Кроме трансформированных клеток, на них реагируют почти все клетки, участвующие в воспалительной реакции. В частности, ФНО усиливают экспрессию молекул адгезии на эндотелии, оказывают хемотаксиче-ское действие на фагоциты, обусловливают синтез белков острой фазы воспаления. В развитии иммунного ответа они участвуют в качестве кофакторов ростовых цитоки-нов, индуцирующих пролиферацию В- и Т-лимфоцитов, усиливают антителообразование, подавляют гиперчувствительность замедленного типа, препятствуют формированию иммунологической толерантности. Большой интерес вызвали данные о том, что ФНО участвуют в морфогенезе лимфоузлов и пейеровых бляшек, а также формировании зародышевых центров при иммунном ответе.
|
|
|
Интерфероны
Согласно принятой номенклатуре выделяют три вида интерферонов - а (макрофагальный), Р (фибробластный), у (лимфоцитарный, иммунный), но по биологическим эффектам их «резистентности» и «Противовирусный иммунитет»), и ИФН II типа, к которому относят γ -интерферон.
ИФН II типа. Источником у-интерферона являются как цитотоксические, так и хелперные лимфоциты, но после дифференциации ТхО на Txl и Тх2 способность вырабатывать его сохраняют только Txl-клетки.
γ -интерферон по структуре представляет собой глико-протеин с молекулярной массой 20-25 кД. После действия антигенов или митогенов пик экспрессии его гена приходится на 48-72 ч. Образование γ -интерферона индуцируется вирусами, но гораздо слабее, чем ИФН I типа. Его продукция в ходе дальнейшего синтеза усиливается и потенцируется ИЛ-2.
По эффектам противовирусного и антиопухолевого действия γ -интерферон уступает α - и β -интерферонам, но намного превосходит их в иммунорегуляции. Он служит стимулятором макрофагов, способствуя обработке и презентации ими антигенов на молекулах МНС класса II, выработке ИЛ-1 и, главное, ИЛ-12, который усиливает его синтез и вызывает дифференциацию Т-хелперов в направлении Txl-клеток, что, в конечном итоге, приводит не просто к снижению секреторной функции Тх2-клеток, а к подавлению гуморального иммунного ответа и развитию клеточного. Являясь продуктом цитотоксических Т- и NK-клеток, у-интерферон, по-видимому, участвует в реализации вызываемого ими цитолиза.
Колониестимулирующие факторы
В группу КСФ входят гранулоцитарно-макрофагоци-тарный (ГМ-), моноцитарно-макрофагальный (М-) и грану лоцитарный (Г-) гемопоэтины. Все эти цитокины представляют собой гликопротеины с молекулярной массой 21-25 кД (ГМ-КСФ), 70-76 кД (М-КСФ), 19-25 кД (Г-КСФ). Образуются клетками стромы, костного мозга и других органов, макрофагами и активированными Т-лим-фоцитами.
Действие КСФ осуществляется в тесной связи друг с другом и с ИЛ-3, 7, 11. Точнее говоря, КСФ поддерживают пролиферацию и начальные этапы дифференциации юных гранулоцитарно-макрофагальных предшественников. В меньшей мере влияют на формирование специализированных моноцитарно-макрофагальных колоний, а также активность зрелых моноцитов, макрофагов и гранул оцитов.
|
|
|
Трансформирующий фактор роста β
ТФРр представляет собой гомодимер (молекулярная масса одной цепи 25 кД). Обладает очень широким спектром действия. В отличие от других факторов роста (в том числе ТФРа) проявляет активность в отношении клеток иммунной системы. Продуцируют ТФРβ огромное количество стромальных клеток и макрофагов, а также клетки многих злокачественных опухолей. Столь же многочисленны его клетки-мишени. ТФРР подавляет синтез многих цитокинов, блокирует ответ лимфоцитов на ИЛ-2, 4 и 7, угнетает формирование NK-клеток и ЦТЛ. Вызывает переключение выработки IgG на IgA. При прекращении выработки ТФРР развивается генерализованный аутоиммунный процесс. 
Теории антителообразования и взаимодействия антител с антигенами
Теория «боковых цепей». Механизм образования антител и их взаимодействие с антигенами впервые обосновал в теории «боковых цепей» П. Эрлих (1898). Представив клетку в виде атомных групп и радикалов, он как химик полагал, что центрально расположенные группировки могут определять ее видовую специфичность, а поверхностные («боковые цепи») - выполнять функцию рецепторов антигенов. Соединяясь с ними, клетка, по мнению П. Эрлиха, затем вырабатывает их так много, что они, не удерживаясь на мембране, отщепляются и поступают в кровь, где функционируют как антитела, способные связывать микробы и токсины. При этом П. Эрлих представлял, что клетка якобы может иметь антитела трех порядков: антитела первого порядка, обусловливающие нейтрализацию токсина, имеют одну группу связывания, которая соединяется с гаптеном антигена; антитела второго порядка, вызывающие реакции агглютинации и преципитации, обладают группой связывания и эффекторной группой склеивания или осаждения антигенов; антитела третьего порядка, связывающие комплемент, содержат две группы связывания: одна вступает в реакцию с антигеном, а другая -с комплементом.
Современные теории антителообразования подразделяют на три группы: 1) инструктивные - теории прямой и непрямой матрицы; 2) селекционные - теории непрямого отбора и селекции клонов антителообразующих клеток; 3) дерепрессии генов.
Теория прямой матрицы. Выдвигая эту теорию в 1930 г., Ф. Брейнль и Ф. Гауровитц предполагали, что синтез антител в клетках происходит на детерминантах антигена, которые, являясь матрицей, определяют структуру и конфигурацию антигенсвязывающих центров (рис. 66, а). Выработанные таким образом под воздействием антигенов иммуноглобулины с центрами связывания в виде слепков детерминант антигена приобретают высокую степень специфичности и способность реагирования лишь с комплементарными гаптенами.
|
|
|
Теория непрямой матрицы. Создатели этой теории Ф. Вернет и Ф. Финнер в целом разделяя идею матричного характера формирования специфичности иммуноглобулинов, в 1949 г. высказали гипотезу опосредованного воздействия антигенов на генотип антителообразующих клеток (рис. 66, б). При этом непрямая информация образования комплементарных антигену иммуноглобулинов передается потомкам клеток.
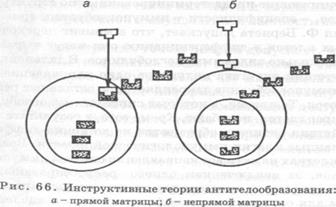 Большой научной ценности теория «боковых цепей» и матричные теории антителообразования в настоящее время не имеют, но на их основе формировались селекционые теории и теория дерепрессии генов.
Большой научной ценности теория «боковых цепей» и матричные теории антителообразования в настоящее время не имеют, но на их основе формировались селекционые теории и теория дерепрессии генов.
Теория непрямого отбора. Выдвинутая в 1955 г. Н. Йерне теория непрямого отбора допускала наличие в организме многочисленных антителопродуцирующих клеток, а в крови - спонтанно выработанного огромного набора антител ко всевозможным антигенам. Антителообразование к каждому из них объяснялось тем, что комплекс антитело - антиген побуждал выработку только таких иммуноглобулинов, которые в нем содержались. Таким образом, антиген в антител ообразовании выполнял роль селектора, а не сигнального индуктора.
Теория селекции клонов антителообразующих клеток. Разработанная Ф. Бернетом в 1959 г. теория селекции клонов антителообразующих клеток постулирует: в процессе длительной эволюции в организме сформировалась гетерогенная популяция антителообразующих клеток, способных продуцировать антитела определенной специфичности; антителообразование детерминировано генотипом и не зависит от антигена; антиген лишь стимулирует (селекционирует) клоны тех антителообразующих клеток, которые имеют к нему специфические рецепторы; в ответ на антигенный стимул происходит пролиферация антителообразующих клеток, а параллельно - и дифференциация предадаптированных клеток в плазмоциты, продуцирующие преддетерминированные по структуре и, главное, - специфичности - иммуноглобулины (рис. 67). Теория Ф. Бернета допускает, что в момент перехода исходных клеток в дифференциацию они могут вырабатывать несколько видов иммуноглобулинов. В дальнейшем в присутствии антигена-индуктора одно направление синтеза иммуноглобулина закрепляется, а остальные репрессируются. Те клетки, в которых синтез иммуноглобулина не закрепляется, погибают. Кроме того, в результате взаимодействия антигена образуются не до конца дифференцированные клетки иммунологической памяти. Возможно, в клетках памяти все направления синтеза иммуноглобулинов, за исключением одного, репрессированы. Повторное внедрение антигена для этих клеток служит не сигналом для осуществления выбора синтеза определенного
 иммуноглобулина, как для исходных клеток, а директивным включателем уже детерминированной дифференциации и пролиферации.
иммуноглобулина, как для исходных клеток, а директивным включателем уже детерминированной дифференциации и пролиферации.
Данная теория охватывает многие стороны иммунитета. Объясняя процесс антителообразования селекцией антигеном соответствующих клонов клеток-предшественников или отбором только некоторого количества из них, она допускает мутационное формирование иммунокомпетентных клеток в онтогенезе. С позиций этой теории стало возможным объяснить состояние толерантности организма к собственным антигенам, под воздействием которых в эмбриональном периоде происходит гибель клеток со специфическими для них рецепторами.
Теория Ф. Бернета стала ключевой и определяющей в развитии современной иммунологии, в частности иммуногенетики.
Теория дерепрессии генов. Теория допускает, что иммунокомпетентная клетка может вырабатывать антитела самой различной специфичности. По минимальным расчетам, ее код содержит информацию, достаточную для формирования 175 ООО антигенсвязывающих центров антител. Однако способность генов детерминировать продукцию такого количества антител репрессирована. Предполагается, что антиген соединяется с ферментом, который обусловливает синтез репрессора и, снимая его действие, индуцирует антителообразование.
Теория взаимодействия антител с антигенами. По вопросу механизма реагирования антигенов с антителами
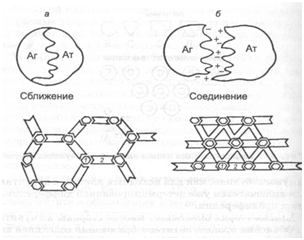 Рис. 68. Взаимодействие антител (Ат) с антигенами (Аг): а - «решетка», б - «каркас»; 1 - Аг; 2 - Ат
Рис. 68. Взаимодействие антител (Ат) с антигенами (Аг): а - «решетка», б - «каркас»; 1 - Аг; 2 - Ат
имеется несколько гипотез. П. Эрлих предполагал, что иммунологические реакции протекают по типу химических и, в частности, представлял, что взаимодействие токсина с антитоксином протекает, как реакция между сильной кислотой и сильной щелочью. Вскоре, однако, выяснилось, что в основе реакции иммунитета лежит адсорбционный процесс, специфичность которого определяется строением детерминантных групп антигенов и соответствующих им по заряду, структуре и конфигурации анти-генсвязывающих центров антител.
Относительно характера соединения антитела с антигеном существуют две гипотезы. По мнению П. Маррека, реакция взаимодействия антигена с антителом обусловлена разноименностью их электрических зарядов (рис. 68). Прочность соединения зависит от количества и полноты совпадения
реагирующих групп антигена и антитела. При оптимальном соотношении антител и антигенов их молекулы в специфических комплексах располагаются в чередующемся порядке и образуют своеобразные «решетки». Л. Полинг рассматривает взаимодействие антитела с антигеном как процесс реагирования двухвалентных и поливалентных структур. Он считает, что при полном насыщении всех валентностей образуются комплексы антигена и антитела в виде «каркаса».
Представленные формы реагирования находят подтверждение при прямых электронно-микроскопических исследованиях различных иммунных реакций, в которых участвуют разнообразные антигены и антитела.
Идиотипы антител как регуляторы иммунного ответа. Вырабатываясь под воздействием антигенов, антитела, связываясь и разрушая их, постепенно ингибируют и прекращают иммунный ответ. Вместе с тем, обладая свойствами антигенов, они, по гипотезе Н. Йерне, могут осуществлять регуляцию иммунитета в сложных сетевых взаимодействиях идиотип-антиидиотип между лимфоцитами различных Т- и В-субпопуляций. Для понимания этого механизма напомним, что каждый специфический антигенсвязывающий фрагмент антитела имеет свой характерный набор идиотипов. В общей массе в организме животных их многие миллионы. Так как огромное разнообразие идиотипов антител, по мнению Н. Йерне, обусловлено многообразием природных антигенных детерминант, то распознающие их лимфоциты должны распознавать также идиотипические детерминанты всех антител, в том числе цитофильных, т. е. самих лимфоцитарных рецепторов. И действительно, так оно и есть. Толерантности к идиотипам у иммунной системы не возникло вследствие ничтожно малых (следовых) их количеств.
 Иммунизированное каким-либо внешним антигеном животное сначала будет вырабатывать против него специфические антитела, затем - антитела к идиотипам антител, а далее - антитела к антиидиотипическим антителам и т. д. Такой цепной тип реакций, дающих «сеть» аутоантител, возникает, когда большая часть выработанных на антигенный стимул иммуноглобулинов имеет одинаковый идиотип. При этом наряду с процессом выработки аутоантител к доминантному идиотипу активируются соответствующие клоны Т-клеток. Те и другие способны ингибировать или усиливать ответ лимфоцитов, имеющих рецепторы к данному идиотипу. Через взаимодействия идиотип-антиидиотип лимфоциты функционально связаны между собой в замкнутую идиотипическую сеть.
Иммунизированное каким-либо внешним антигеном животное сначала будет вырабатывать против него специфические антитела, затем - антитела к идиотипам антител, а далее - антитела к антиидиотипическим антителам и т. д. Такой цепной тип реакций, дающих «сеть» аутоантител, возникает, когда большая часть выработанных на антигенный стимул иммуноглобулинов имеет одинаковый идиотип. При этом наряду с процессом выработки аутоантител к доминантному идиотипу активируются соответствующие клоны Т-клеток. Те и другие способны ингибировать или усиливать ответ лимфоцитов, имеющих рецепторы к данному идиотипу. Через взаимодействия идиотип-антиидиотип лимфоциты функционально связаны между собой в замкнутую идиотипическую сеть.
|
|
|
