 |


Сегментарные механизмы формирования болевого потока и восходящие пути болевой чувствительности
|
|
|
|
Первым центральным звеном, воспринимающим разномодальную афферентную информацию, является нейрональная система заднего рога спинного мозга. По современным данным, она представляет собой цитоархитектонически весьма сложную структуру, а в функциональном отношении рассматривается как своеобразный первичный интегративный центр сенсорной информации. Именно здесь происходит взаимодействие между каналами болевой и неболевой чувствительности, на основании которого формируется поток восходящей импульсации нового качества. В настоящее время в мировой литературе имеется огромное количество публикаций, посвященных морфофункциональной организации этой области спинного мозга и ее роли в переработке, формировании и передаче ноцицептивной информации, которые обобщены в ряде монографий и обзоров [Костюк П.Г., Преображенский Н.Н., 1975; Вальдман А.В., Игнатов Ю.Д., 1976; Жукова Г. П., 1977; Brown Н., 1981; Dubner R. et aL, 1984; Peri E., 1984; Lggo A. et al., 1985; Sigiura Y. et al., 1986; Light A., 1988; Blumenkopf В., 1988].
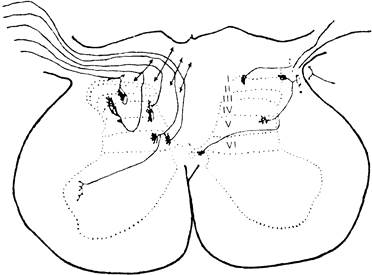
Рис 1 Распределение окончаний первичных афферентов в спинном мозге (по A. Light, 1988).
Слева - окончания афферентных волокон разных типов от сухожильных рецепторов, мышечных веретен и кожи, не связанных с ноцицепцией; справа - окончания безмиелиноных С афферентов и тонких миелиновых волокон, связанных с ноцицепцией, I — VI — слои серого вещества спинного мозга по В. Rexed (1954).
Распределение первичных афферентов в заднем роге спинного мозга. Афферентные волокна различного калибра по-разному распределяются уже в задних корешках перед их входом в спинной мозг — толстые миелинизированные проводники занимают их медиальную часть, а тонкие миелинизированные и С-волокна располагаются более латерально. Перерезка этих латеральных пучков сопровождается уменьшением боли. После входа в спинной мозг А-δ- и С-волокна в составе тракта Лиссауэра идут в каудальном и ростральном направлениях в пределах 1 — 2 сегментов и оканчиваются в дорсальной части заднего рога (рис. 1). Имеется определенное различие в окончании А-δ-, С-волокон, передающих ноцицептивную информацию, и А-δ-волокон, связанных с механорецепторами.
|
|
|
Установлено, что в I пластине, содержащей крупные, маргинальные клетки, оканчиваются А-δ-волокна, активируемые термическим и механическим ноцицептивными раздражениями, и С-полимодальные ноцицентивные афференты. Широко ветвящиеся тонкие терминали С-волокон образуют синапсы на дендритах маргинальных клеток и аксоаксональные контакты на терминалях других афферентных волокон. Желатинозная субстанция (II пластина) является основным местом окончания С-волокон, а также других афферентов, которые входят в нее с дорсальной поверхности, и, широко разветвляясь, диффузно распределяются во всей ее толще, достигая вентральной границы. А-δ-волокпа также оканчиваются в желатинозной субстанции. Причем А-δ-афференты, связанные с механорецепторами, оканчиваются в центральной части II пластины и в III пластине и более вентрально, тогда как ноцицентивные А-δ-волокна распределяются в верхних отделах II пластины.
Толстые афферентные волокна после входа в спинной мозг идут в составе дорсальных столбов и отдают коллатерали, оканчивающиеся на нейронах IV — VI пластин заднего рога. Некоторые волокна, делая в этой области восходящую петлю, входят в желатинозную субстанцию с вентральной стороны и разветвляются в радиальном направлении по всей ее глубине. Терминали этих волокон образуют синапсы на нейронах желатинозной субстанции и на дендритах глубжерасположенных клеток. В настоящее время очень мало известно о нейрохимической медиации в самом первом центральном синапсе: афферентное волокно-нейрон заднего рога спинного мозга. Предполагается несколько кандидатов, наиболее вероятными из которых считаются субстанция Р, холецистокинин, соматостатин, вазоактивный интестинальный полипептид (ВИП) [Dodd J. et al., 1984; Bhnnenkopf В., 1988].
|
|
|
Функциональная характеристика нейронов заднего рога, связанных с ноцицептивной афферентацией. Все нейроны дорсальной части заднего рога могут быть разделены на 3 группы. Первая состоит из клеток, активируемых исключительно ноцицептивными стимулами или раздражением «ноцицентивных» А-δ- и С-волокон. Вторая группа включает клетки, отвечающие на ноцицептивные и неноцицептнвные воздействия и на стимуляцию как высокопороговых, так и низкопороговых афферентов. Третья группа представлена нейронами желатинозной субстанции, которые, взаимодействуя с клетками первых двух групп, образуют систему модулирующего контроля афферентного входа в спинной мозг и формирования восходящего ноцицептивного потока импульсов.
Нейроны, активируемые ноцицептивными стимулами, так называемые специфические ноцицептивные нейроны локализованы в основном в I слое, где оканчиваются А-δ- и С-афференты. Они имеют ограниченные (3 см2) рецептивные поля и небольшую пространственную конвергенцию афферентных входов. Некоторые из них, как правило, не имеющие спонтанных разрядов, активируются только А-δ-волокнами, связанные с механорецепторами. Другие, спонтанно активные, отвечают на механическое, термическое ноцицептивное раздражение кожи и на электрическую стимуляцию А-δ- и С-волокон (рис. 2), причем возбуждающее действие С-волокон на эти клетки не зависит от соотношения активности в толстых и тонких немиелинизированных волокнах, так как ответ нейронов на тепловое ноцицептивное раздражение кожи очень мало изменялся при блоке проведения по А-волокнам.

Рис. 2. Ответ нейрона заднего рога спинного мозга на электрораздражение С-афферентов до (а), после (б) поляризации афферентных волокон и локализация нейрона (в).
Сверху вниз: С-ответ кожного нерва при сверхпороговом раздражении, ответ нейрона на раздражение С-афферентов.
|
|
|
По некоторым данным, более 25 % нейронов желатинозной субстанции также реагировали исключительно на ноцицептивные стимулы. Предполагается, что такие клетки, активируемые С-волокнами, в функциональном отношении являются вставочными нейронами на пути передачи импульсов от первичных афферентов к нейронам I пластины. При одновременной регистрации активности пары клеток — одной в желатинозной субстанции, другой — в I пластине — было установлено, что латентный период первого потенциала действия в ответе нейрона желатинозной субстанции меньше, чем латентный период I спайка в ответной реакции нейрона I пластины и оба этих нейрона имели перекрывающиеся рецептивные поля [Price D. et al., 1979]. Эти различия были особенно характерны для нейронов наружной зоны желатинозной субстанции. Именно здесь располагаются клетки стеблевидной формы с аксонами, направляющимися в I пластину заднего рога. Кроме того, клетки желатинозной субстанции, активируемые С-афферентами, могут возбуждать нейроны не только I, но и V пластины. Различное региональное распределение клеток I пластины, которые получают ноцицептивный вход через А-δ-афференты, и клеток желатинозной субстанции, возбуждаемых С-волокиами, могут быть одной из причин возникновения «быстрой» и «медленной» боли, поскольку аксоны нейронов I пластины участвуют в формировании спиноталамической восходящей системы.
Предполагается, что «специфические» ноципептивные нейроны играют существенную роль в формировании восходящего ноцицептивного потока, который передает в высшие отделы мозга информацию об интенсивности и локализации ноцицептивных стимулов (более подробно о морфологии и свойствах этих нейронов см.: Игнатов Ю.Д., 1986; Diidner R., Bennett G. 1983; Rethelyi M. et al, 1983; Woolf С., Fitzgerald M, 1983 Peri E., 1984; Iggo A. et al., 1985).
Однако ряд экспериментальных данных показывает, что роль клеток I слоя заднего рога, активируемых исключительно ноцицептивными афферентами, в общей картине процессов формирования восходящего болевого потока не должна излишне переоцениваться, как это было сразу после выявления этих «специфических» ноцицептивных нейронов Оказалось, что таких «специфических» нейронов не так уже много в I пластине и что они перемешаны с клетками, отвечающими на стимуляцию как высоко-, так и низкопороговых афферентов, а некоторые из них активируются и раздражением А β-афферентов и даже мы щечных волокон III и IV групп [Price D. et al.. 1979]. Кроме того, эти клетки не активировались брадикинином, и их ответы на ноцицептивныс воздействия не коррелировали с поведенческим реагированием на боль. Выявлено, что аксоны нейронов I пластины проецируются не только в спиноталамический тракт, но и во все квадранты спинного мозга и даже в такой классически неболевой тракт, как дорсолатеральные канатики [МсМаhan S., Wall P., 1982].
|
|
|
Нейроны, активируемые разномодальными афферентными входами, или «мультирецептивные», «конвергентные», «нейроны широкого афферентного диапазона», локализованные в основании заднего рога (пластины IV — V) и глубже (частично пластины VI), играют основную роль в переработке ноцицептивной информации, в передаче ее в восходящем направлении и в реализации низкоинтегрированных простейших реакций на повреждающие воздействия. Строение, источники их активации, свойства этих клеток и проекции их аксонов хорошо исследованы [Brown А., 1982; Dubner R., Bennett О., 1983; Peri E., 1984; Maixner W. et al, 1986; Li^ht A, 1988].
Общепризнанно, что клетки основания заднего рога имеют достаточно широкое, хорошо контурируемое рецептивное поле, окруженное подпороговой зоной, имеющей незначительный афферентный вход на эти нейроны. В тех случаях, когда эффективность афферентного входа возрастает, рецептивное поле может увеличиваться [Wall Р, Devor M., 1981]. На этих нейронах конвергируют афферентные волокна различного спектра и модальности, поскольку они отвечают на прикосновение, давление, на механическую и термическую ноцицептивную стимуляцию рецептивного поля. Они моносинаптически активируются низкопороговымн А-волокнами и одновременно имеют полней наптический С-вход. Нейроны IV—V пластин отвечают высокочастотной «пачкой» импульсов на естественное неповреждающее раздражение кожи и длительной импульсной активностью при ноцицептинном воздействии. Частота и продолжительность ответов нейрона возрастает пропорционально интенсивности ноципептивных, особенно термических стимулов.
Нейроны V пластины реагируют с латентным периодом 15—30 мс длительной и высокочастотной (частота разрядов увеличивается на 100—800%) импульсной активностью на внутриартериальное введение брадикинина, причем латентный период и продолжительность ответной реакции на брадикинин у приматов хорошо коррелируют с возникновением ощущения боли у человека.
|
|
|
Одновременное раздражение А- и С-афферентов проявляется двумя ответными реакциями клеток — ранней (коротколатентный ответ на стимуляцию А-волокон) и поздней (С-ответ). Выключение проведения возбуждения по А-волокнам их поляризацией усиливает ответы клеток на раздражение С-волокон, и наоборот, предшествующее раздражение А-афферентов уменьшает их реагирование на ноцицептивное раздражение рецептивного поля или на активацию С-волокон. Нами было обнаружено, что ответная реакция нейронов V слоя на ноцицептивное механическое воздействие уменьшается, если оно наносится на фоне (или сразу же после) ритмической стимуляции низкопороговых афферентов (рис. 3). В последнее время получены весьма убедительные доказательства ведущей роли конвергентных нейронов в процессах кодирования интенсивности ноцицептивных термических стимулов [Maixner W. et al., 1986]. Предполагается, что изменение свойств конвергентных нейронов: рецептивного поля, спонтанной активности и ответов на разномодальные раздражения — у крыс с экспериментальным артритом лежат в основе гипералгезии и болевого синдрома у больных ревматоидным артритом [Caivino В. et al., 1987].
На нейронах заднего рога конвергируют не только разномодальные кожные, но также мышечные (III—IV группа) и висцеральные афференты. Происходящее на этих нейронах взаимодействие соматовисцеральных афферентных потоков лежит в основе возникновения отраженной боли [Foreman R. et al., 1981]. Установлено, что нейроны, на которых осуществляется взаимодействие соматических и висцеральных афферентов, активируются через висцеральные входы при естественном раздражении висцеральных рецепторов, но только в том случае, когда интенсивность воздействия, например давление в желчных путях достигает ноцицептивного уровня [Cervero К, 1982, 1985].

Рис. 3. Ответ нейрона IV слоя заднего рога спинного мозга на ноцицептивное раздражение кожи, наносимое до и после ритмической стимуляции пизкопороговых афферентов.
1 — фоновая импульсная активность: 2 — импульсная активность при ноцицептивном раздражении до стимуляции низкопороговых афферентов: 3 — то же после ритмического раздражения низкопороговых афферентов. Линия под осциллограммами — период ноцицсптпвного воздействия на кожу. Калибровка: времени — 1 с, амплитуды — 250 мкВ.
Естественно, что не все нейроны заднего рога имеют ноцицептивные афферентные входы. Во всех пластинах, особенно в IV, имеется достаточно большая популяция клеток с узким функциональным диапазоном, которые активируются легкими механическими воздействиями (прикосновением, небольшим давлением) и не отвечают на повреждающие стимулы. В то же время клетки, отвечающие на ноцицепцию, выявлены в VI пластине и даже в вентральном роге [Molinary H., 1982]. Эти нейроны, вероятно, участвуют в регуляции не только моторной, но и сенсорной деятельности спинного мозга, и в частности в регуляции боли, способствуя локализации ответа на боль. Поэтому цитоархитектонически выделяемые пластины в заднем роге спинного мозга скорее должны рассматриваться как зоны преимущественной концентрации нейронов с определенными свойствами, а не как строго функционально специализированные нейронные ансамбли.
Таким образом, можно считать, что конвергентные нейроны, локализованные в основании заднего рога, большинство из которых имеют восходящие проекции, принимают самое непосредственное участие в возникновении информации такого качества, которая высшими отделами головного мозга расценивается как болевая и которая запускает сложные механизмы ответного реагирования на боль. Однако в настоящее время есть все основания считать, что активность релейных нейронов, связанных с ноцицептивной афферентацией, их ответы на разномодальные стимулы, взаимодействие на них различных афферентных входов и, следовательно, формирование восходящего импульсного потока модулируется нейронами желатинозной субстанции.
Нейроны желатинозной субстанции, локализованные во II пластине, стали предметом активного исследования в последние 15—20 лет. Морфофункциональная организация этой области детально рассматривается в специальных обобщающих статьях и обзорах [Bennett G. el al., 1980; Cervero F., I^go A., 1980; Wall P., 1980; Rethelyi M. et al., 1982; Dubner R, Bennett G., 1983; Bicknell Jr., Beal J., 1984; Dubner R. et al., 1984; Per! E, 1984; Iggo A. et al., 1985]. Поэтому здесь мы коснемся лишь некоторых свойств нейронов желатинозной субстанции, необходимых для понимания их роли в регуляции ноцицепции.
Эти нейроны имеют рецептивные поля различных размеров, формы и локализации. У половины из них рецептивные поля по размерам и локализации соответствуют таковым у нейронов IV—V пластин и располагаются в пределах какой-либо одной области (стопа, пальцы) нижней конечности. Характерной особенностью 1/3 нейронов желатинозной субстанции является наличие у них очень маленьких (менее 2 см2) рецептивных полей [Wall P., 1980]. У незначительного количества (3—9%) нейронов рецептивные поля занимали всю поверхность ипсилатеральной и иногда даже контралатеральной конечности. Именно на них конвергируют А-δ-, С-волокна, а на некоторых и А-р-афференты. Примечательно, что аксоны этих клеток не выходят за пределы спинного мозга.
Ориентация аксонов желатинозных нейронов весьма различна и во многом определяется локализацией сомы клетки. У нейронов, расположенных в дорсальной части желатинозной субстанции, аксоны направлены дорсально и проецируются в тракт Лиссауэра. Аксоны нейронов средней трети идут в продольном направлении внутри желатинозного вещества. По мере приближения к нижней границе желатинозной субстанции аксоны принимают вентральное направление и проникают, как правило, в глубжерасположенные слои заднего рога. О протяженности аксональных проекций имеются разноречивые и даже полярные представления. Согласно одним данным, аксоны нейронов желатинозной субстанции оканчиваются в пределах одного сегмента [Beal J., Cooper M., 1978], в то время как по другим — они достигают даже таламуса [Willis W. et al., 1978]. Однако аксоны большинства (до 80%) желатинозных нейронов не проецируются дальше одного сегмента и оканчиваются в пластинах I, IV, V или даже в пределах одной II пластины. У меньшей части нейронов аксоны, входящие в тракт Лиссауэра, прослеживаются на расстоянии 15 — 40 мм.
Отдельные клетки имеют контралатеральные аксональные проекции.
Основная масса нейронов имеет спонтанную активность в диапазоне от 1 до 40 имп/с (в среднем 5 — 10 имп/с). Она представлена разрядами постоянного ритма без высокочастотных «пачек» импульсов. Ни перерезка дорсолатеральных канатиков, ни полная спинализация существенно не изменяли фоновую активность желатинозных нейронов, что свидетельствует о ее независимости от нисходящей тонической регуляции и о большой роли афферентного сегментарного входа в ее формировании [Steedman W., Molony V., 1983]. Нейроны желатинозной субстанции реагируют на естественное раздражение их рецептивного поля. Большинство клеток отвечали как на неповреждающую механическую (прикосновение, расчесывание), так и на ноцицептивную (сдавленно зубчатым зажимом) стимуляцию [Price D. et al., 1979; Fitzgerald M., 1981]. По данным Р. Cervero и A. Iggo (1980), разномодальная активация рецептивного поля сопровождается исключительно торможением активности нейронов, и в зависимости от характера воздействия они могут быть разделены на 3 группы. В первую группу входят клетки, активность которых тормозилась при низкопороговой механической стимуляции рецептивного поля. Вторую группу составляют нейроны, торможение активности которых возникало при раздражении рецептивного поля как повреждающими, так и неповреждающими стимулами. К третьей группе были отнесены клетки, разряды которых угнетались только при ноцицептивном раздражении рецептивного поля. Эти 3 группы клеток в функциональном отношении, т.е. по характеру ответов на разномодальную стимуляцию рецептивного поля, являются антиподами глубжерасположенных нейронов.
По общепринятому мнению, клетки желатинозной субстанции отличаются от других нейронов заднего рога, в частности V пластины, характером ответов на раздражение С-волокон и взаимодействием А- и С-афферентных входов. Так, нейронам желатинозной субстанции не свойствен феномен «взвинчивания» (wind up). Они не изменяют, как нейроны V пластины, ответы на раздражение С-волокон при выключении А-афферентов, а предшествующая стимуляция их не уменьшает ответные реакции желатинозных нейронов, возникающие при внутриартериальном введении брадикинина [Fitzgerald M., 1981]. Наряду с хорошо изученным быстрым реагированием нейронов желатинозной субстанции на разномодальные афферентные воздействия, выявлены более медленно и длительно протекающие изменения их активности [Wall P., 1980, 1985]. Эти изменения, вероятно, обусловлены действием различных биологически активных соединений, осуществляющих нейромодуляторную функцию, таких как энкефалпны, соматостатин, ангиотензин, холецистокинин, гликокортикоиды, субстанция Р, ГАМ.К, норадреналин, серотонин, локализующихся в дорсальной части заднего спинного мозга [Hunt S., 1982; Dodd J. et al., 1984; Blumenkopf В., 1988]. Однако функциональная роль медленных, длительных изменений активности нейронов желатинозной субстанции и их связь с регуляцией ноцицептивного афферентного потока пока не выяснены и требуют новой концептуальной гипотезы.
Функциональная роль желатинозной субстанции до настоящего времени весьма спорна. Практически все исследователи придают ей важное значение как нейрональной системе, регулирующей сенсорный вход и болевую чувствительность на сегментарном уровне. Однако каждый имеет свое, порой весьма противоречащее имеющимся, представление о механизмах этой регуляции. В настоящее время снова получила подтверждение и развитие точка зрения, высказанная более 30 лет назад [Pearson Л., 1952], о том, что желатинозная субстанция функционирует как релейная инстанция, передающая ноцицептивную информацию от первичных афферентов к нейронам спиноталамического тракта. Именно в области желатинозной субстанции оканчивается основная масса ноцицептивных А-δ- и С-афферентов, которые непосредственно активируют нейроны желатинозной субстанции [Kumazawa Т., Peri Б., 1978; Peri Б., 1984].
Согласно другой, более общепринятой концепции клетки желатинозной субстанции не просто передают, а модулируют, эффективность синаптического воздействия афферентных входов на нейроны дорсального рога спинного мозга. Традиционно считается, что эта модуляция реализуется тормозными механизмами и что большая часть клеток желатинозной субстанции функционирует как тормозные интернейроны. Примечательно, что налоксон при внутривенном введении, возбуждая нейроны V пластины, одновременно угнетает активность нейронов желатинозной субстанции [Fitzgerald M., Wall P., 1980].
Модуляция афферентного сегментарного входа клетками желатинозной субстанции может осуществляться как пост-, так и пресинаптическими механизмами. В настоящее время известно немало достаточно хорошо проверенных фактов, свидетельствующих о важной роли желатинозной субстанции в генерации деполяризации первичных афферентов (ДПА) [подробнее см.: Wall M., 1980]. Желатинозной субстанции отводится ключевая роль в одной из самых распространенных и, пожалуй, самых дискуссионных теорий боли — теории «входных ворот» (gate control theory of pain) P. Мелзака и Р. Уолла (1965) [подробнее см. Мелзак Р., 1981]. Эта теория в первом ее постулате объясняет формирование ноцицептивного потока на сегментарном уровне на основе центрального взаимодействия быстропроводящей миелиновой (неноцицептивной) и медленнопроводящей безмиелиновой систем на первых релейных (конвергентных, мультирецептивных) нейронах спинного мозга (рис. 4). Афферентная импульсация, возникающая в миелиновых волокнах при неповреждающих раздражениях, одновременно с активацией релейных нейронов (Т) через коллатерали возбуждает нейроны желатинозной субстанции. Они, в свою очередь, деполяризуют первичные миелиновые афференты и тем самым пресинаптически ограничивают активирующее их влияние на Т-клетки. При ноцицептивном воздействии активируются тонкие миелиновые и безмиелиновые волокна, которые (в оригинале теории только С-волокна) оказывают тормозное влияние на нейроны желатинозной субстанции, в результате чего уменьшается деполяризация (или возникает гиперполяризация) миелиновых волокон, т.е. «открывается» афферентный вход и возрастает эффективность синаптического воздействия на Т-нейроны. Т-нейроны при достижении критического уровня, определяемого временной и пространственной суммацией «ноцицептивных» и неноцицептивных сигналов, формируют восходящий поток импульсов нового качества, который воспринимается вышерасположенными структурами мозга.

Рис. 4. Схема системы контроля афферентного входа (по R. Melzack, Р. Wall, 1965).
1 – толстые миелиновые волокна; 2 — тонкие безмиелиновые волокна: SG — желатинозная субстанция; Т — нейроны первого центрального переключения; (+) — возбуждающее действие. (—) — тормозное действие.
Положение теории Р. Мелзака и П. Уолла о том, что процессы пресинаптического торможения и облегчения являются ведущими в реализации модулирующего влияния желатинозной субстанции, длительное время являлись предметом активного обсуждения. Эта теория, как и любая новая теория, постоянно совершенствуется и дополняется. В современных ее модификациях, и в том числе самих авторов [Vyklicky L., 1981; Wall P., 1978, 1984], больший акцент сделан на роль постсинаптических и нейрохимических процессов в регуляции передачи разномодальных импульсов на релейные нейроны спинного мозга.

Рис. 5. Схема лемнисковой и экстралемнисковой восходящих афферентных систем (по Р. Boulu, 1984).
FST — спиноталамический тракт, FSRT — спиноретику-лоталамический тракт.
Таким образом, можно считать, что формирование восходящего «ноцицептивного» потока в спинном мозге происходит двумя способами — на основании взаимодействия разномодальных афферентных входов на релейных (конвергентных) нейронах и в результате активации «специфических болевых» нейронов, имеющих восходящие проекции.
Восходящие пути болевой чувствительности. Существуют 2 основные «классические» — лемнисковые и экстралемнисковые — восходящие афферентные системы (рис. 5). В пределах спинного мозга одна из них располагается в дорсальной и дорсолатеральной зоне белого вещества, другая — в его вентролатеральной (антеролатеральной) части. Известно, что в ЦНС не существует специализированных путей болевой чувствительности и что интеграция боли осуществляется на различных уровнях ЦНС на основе сложного взаимодействия лемнисковых и экстралемнисковых проекций. Вместе с тем не вызывает сомнений тот факт, что значительно большую роль в передаче восходящей ноцицептивной информации играют вентролатеральные проекции, морфофункциональная организация которых детально исследована и обобщена в ряде монографий и обзоров [Оганнсян А.А., 1978; Yaksh Т., Hammond D, 1982: Kevetter G, Willis W., 1983; Ralston H., 1984; Willis W., 1985]. Об этом свидетельствуют и клинические наблюдения, показывающие, что после антеролатеральной хордотомии или деструкции этой системы при некоторых заболеваниях возникает длительная и выраженная аналгезия. С другой стороны, ощущение боли и ее локализация сохраняются при перерезке всех, кроме вентролатеральных, путей, а стимуляция аксонов в антеролатеральном квадранте после хордотомии вызывает интенсивную боль [Noordenbos W„ Wall P., 1976].
Источниками вентролатеральных проекций являются нейроны I—V и даже VI—VIII пластин спинного мозга (рис. 6), аксоны которых имеют диаметр от 1 до 11 мкм и скорость проведения 7—74 м/с. Вентролатеральные восходящие пути в пределах спинного мозга имеют определенную соматотопическую организацию — волокна, исходящие из более каудальных сегментов, располагаются более латерально, а из ростральных — медиально и вентрально. Конечные проекции первоначально выявляются в продолговатом мозге, затем — в среднем и, наконец, в галамусе. В зависимости от места окончания восходящая антеролатеральная система разделяется на 3 основных тракта — спиноталамический, спиноретикулярный и спиномезенцефалический. Первый из них обозначается как неоспиноталамический, а два других объединяются в палеоспиноталамический тракт. Однако данные последних лет со все большей убедительностью свидетельствуют об относительности такого строгого разделения, поскольку коллатерали аксонов одних и тех же нейронов спинного мозга могут проецироваться в различные супрасегментарные структуры [Kevetter G., Willis W., 1983; Liu R., 1986; Pechura С., Liu R., 1986]. Более того, предполагается, что некоторые свойства нейронов — источников восходящих трактов: размеры рецептивных полей, ответы на разномодальные стимулы, скорость проведения по аксону — различаются в зависимости от проекции их аксонов [Yezierski R. et al., 1988].
Неоспиноталамический тракт начинается от нейронов I, V и частично VI — VII пластин спинного мозга. Нейроны — источники этого тракта, имеют кожные рецептивные поля различных(как правило, среднего и большого) размеров. В зависимости от модальности афферентных входов они разделяются на несколько групп. Одни из них, локализованные преимущественно в I пластине, активируются только ноцицептивными стимулами, другие, в основном клетки IV — V пластин, отвечают и на неповреждающие воздействия. На большинстве нейронов спиноталамического тракта, располагающихся в V пластине, конвергируют мышечные, кожные, висцеральные афференты, и эти клетки отвечают прогрессивно увеличивающейся частотой разрядов на неноцицептивную и разномодальную ноцицептивную (механическую, термическую, химическую) стимуляцию [Milne R. et al., 1981]. Аксоны спиноталамических нейронов после перекреста в передней комиссуре восходят контралатерально в вентролатеральном квадранте спинного мозга (рис. 7). В стволе головного мозга Неоспиноталамический тракт располагается дорсолатеральнее пирамид и нижнеоливарного комплекса и, не прерываясь, достигает таламуса. В пределах заднего таламуса он разделяется на латеральную и медиальную части. Первая из них оканчивается преимущественно в n. ventralis posterior lateralis, а вторая — в медиальных и интраламинарных ядрах. Недавно установлено, что нейроны — источники спиноталамического тракта содержат глутамат и холецистокинин [Ju G. et al., 1987; Magnusson K. et al., 1987]. Выявлено даже различие в их нейрохимической организации в зависимости от проекции аксонов — нейроны, аксоны которых оканчиваются в медиальном таламусе, содержат энкефалин и динорфин, в то время как клетки, проецирующиеся в латеральный таламус, — вазоактивный интестинальный пептид [Nahin R, 1988].

Рис. 6. Схема источников путей, восходящих в антеролатеральных столбах спинного мозга. I — VIII — слои серого вещества спинного мозга по В. Rexed (1954).

|
Рис. 7. Схема спиноталамического тракта (по W. Willis, 1985). Ядра таламуса, с. 1. — центральное латеральное, с. т. — центральное медиальное; 1. р. — латеральное заднее, m. d. — медиальное заднее, р. f. — парафасцикулярное, v, р. I. каудальная часть вентрального заднелатерадь-ного; v. р. m. — вентральное заднемедиальное.
Прецизионными электрофизиологическими и гистологическими методами установлены различные проекции спиноталамических нейронов в таламусе. Аксоны спиноталамических нейронов I пластины и IV—V пластин оканчиваются соответственно в его вентролатеральных и задних ядрах, а глубжерасположенные клетки — в интраламинарных ядрах. Однако тонкая структура терминалей спиноталамических проекций из I и V слоев заднего рога спинного мозга в вентробазальный комплекс таламуса не отличается от таковой у терминалей спиноталамических аксонов клеток VII слоя заднего рога, оканчивающихся в центральном латеральном ядре таламуса [Peschanski M. et al., 1985- Ма W. et al., 1987).
Спиноретикулярный тракт является вторым по величине путем передачи ноцицептивной информации. Нейроны — источники этого тракта, локализуются в V—VII и частично в VIII пластинах спинного мозга, что неоднократно было показано методом антидромной активации и маркировкой этих нейронов с помощью пероксидазы хрена. Некоторые спиноретикулярные нейроны являются энкефалинсодержащими. Примечательно, что нейроны I пластины спинного мозга не участвуют в формировании этого тракта [Kevetter G, Willis W., 1983; Nahin R„ Nicevich P., 1986]. Спиноретикулярные нейроны имеют небольшие кожные рецептивные поля и активируются как неноцицептивными, так и ноцицептивными стимулами, причем частота их разрядов увеличивается с усилением интенсивности раздражения.

Рис. 8. Схема спиноретикулярного тракта (по W. Willis, 1985). n. g. с. — гигантоклеточное ядро.
Рис. 9. Схема спиномезенцефалического тракта (по W. Willis, 1985). β- а. g. — околоводопроводное серое вещество; с. u. n. — клиновидное ядро
Спиноретикулярный тракт оканчивается в каудальной части ствола мозга (рис. 8). Именно в этой области, особенно в ретикулярном гигантоклеточном ядре, парагигантоклеточном ядре и в большом ядре шва, обнаружена массивная конечная дегенерация при перерезке вентролятеральных столбов. У человека больше половины волокон, восходящих в вентролатеральных столбах, оканчивается ипси- и контралатерально в зоне медиальных и латеральных ядер ретикулярной формации продолговатого мозга. Восходящие проекции от этих ядер достигают интраламинарных ядер таламуса (ретикулобульботаламический путь) и диффузно распространяются в гипоталамусе и лимбических структурах.
Спиномезенцефалический тракт иногда рассматривается как компонент спинорстикулоталамической системы, поскольку он формируется аксонами нейронов тех же пластин, где локализованы спиноретикулярные клетки [Menetrey D. et al., 1982]. Однако есть отличие в его морфологической организации и проекциях. Волокна спиномезенцефалического тракта в вентро-латеральных столбах располагаются более поверхностно. Часть из них перекрещивается в спинном мозге, другие восходят преимущественно ипсилатерально и перекрещиваются в области покрышки головного мозга (рис. 9). У кошек часть спиномезенцефалических проекций от нейронов I слоя располагается в дорсолатеральном канатике [Hylnen J. et al., 1986]. Оканчивается спиномезенцефалический тракт в ретикулярной формации среднего мозга в центральном сером веществе и в таламусе. Клетки — источники спиномезенцефалического тракта различаются по локализации в спинном мозге, по размерам рецептивных полей, по ответам на разномодальные стимулы в зависимости от окончания их аксонов [Yezierski R., Schwartz R., 1986]. К этим восходящим волокнам добавляются многочисленные проекции из бульбарных структур, и вся эта сложная система часто объединяется в «диффузный покрышковый» или спинотектальный путь. В среднем мозге он отдает коллатерали в центральное серое вещество и в глубокие отделы верхних бугров. Волокна из центрального серого вещества, в свою очередь, проецируются в медиальные таламические ядра, в вентральную тегментальную область и в гипоталамус.
Антеролатеральные тракты являются основной, но не единственной ноцицептивной системой. Известно, что после антеро-латеральной хордотомии примерно у 50 % лиц через несколько месяцев снова возникала боль. Имеются также многочисленные экспериментальные и клинические данные о невозможности получения стойкой аналгезии при разрушении таламических сенсорных ядер [Tasker R., 1984]. Определенное значение в передаче болевой информации в ростральном направлении имеют тракты, восходящие в дорсальных и дорсолатеральных столбах белого вещества спинного мозга, традиционно относящиеся к лемнисковой афферентной системе.
Спиноцервикальный тракт — отчетливо выражен у низших млекопитающих, особенно кошек, и в меньшей степени у обезьян и человека. Он формируется аксонами клеток IV—VII пластин, которые восходят в дорсолатеральных столбах и оканчиваются в латеральном цервикальном ядре на уровне Ci-ii шейных сегментов. Нейроны — источники этого тракта активируются тактильными и различными (термическим, механическим, химическим) ноцицептивными раздражениями их рецептивных полей [Brown A., 1981]. От латерального шейного ядра афферентные проекции восходят через латеральную часть медиального лемниска, оканчиваются в среднем мозге и в контралатеральном задневентральном ядре таламуса и, переключаясь в нем, достигают соматосенсорной зоны коры. Нейроны латерального шейного ядра активируются также ноцицептивными механическими и термическими стимулами [Kalander К., Giesler G., 1987].
Дорсальные тракты содержат миелинизированные афференты задних корешков от кожи, сухожилий, мышц и внутренних органов, которые без перерыва в сером веществе спинного мозга восходят в задних столбах до нежного и клиновидного ядер. Вторичные проекции от задних ядер столбов через медиальную петлю достигают задневентрального ядра таламуса, а оттуда проецируются в соматосенсорные зоны коры. Одновременно часть коллатералей заднекорешковых волокон оканчивается на нейронах III—IV пластин заднего рога спинного мозга, аксоны которых образуют так называемый постсинаптический дорсальный тракт, который также оканчивается в нежном и клиновидном ядрах [Bennett G. et al., 1983]. Часть
|
|
|
