 |


Нейрофизиологические и нейрохимические механизмы формирования гемодинамических ноцицептивных реакций
|
|
|
|
Боль, выполняющая на начальном этапе несомненно сигнальную функцию, определяет ответные реакции организма, направленные на мобилизацию разномодальных регуляторных систем для устранения ноцицептивного воздействия или его последствий. Наряду с включением механизмов, определяющих эмоционально-аффективные проявления боли, которые в функциональном плане являются одновременно стимулом к активному предотвращению чрезвычайного вредоносного раздражителя, также активируются процессы вегетативного реагирования. Особую значимость среди них имеют изменения сердечно-сосудистой системы. Гемодинамические реакции, возникающие на этапе ощущения боли, «боли-сигнала», направлены прежде всего на обеспечение возросших нутритивных потребностей скелетной мускулатуры, необходимых для реализации комплекса аффективно-моторных актов активной обороны или избегания. С общебиологических позиций такие реакции сердечно-сосудистой системы имеют адаптивное значение.
Иная ситуация складывается при продолжающемся и (или) усиливающемся ноцицептивном воздействии, особенно когда в силу ограничивающих влияний различных факторов (биологических и социальных у человека) гемодинамические сдвиги, запускаемые болевыми стимулами, не могут реализоваться в процессе двигательных актов. В этих условиях сердечно-сосудистые реакции утрачивают свой адаптивный смысл и приобретают значение самостоятельных патогенетических факторов развития патологического состояния. Следовательно, нормализация сдвигов сердечно-сосудистой системы должна быть неотъемлемым компонентом оптимального лекарственного обезболивания. К сожалению, данные экспериментальной фармакологии и опыт клинической медицины свидетельствуют о малой эффективности влияния существующих болеутоляющих средств на гемодинамические реакции ноцицептивного генеза, что во многом обусловлено недостаточной изученностью механизмов регуляции функции сердечно-сосудистой системы при боли. Ключевое значение в этом плане имеет вопрос об общности или разобщенности нейрофизиологических механизмов формирования эмоционально-аффективных и гемодинамических проявлений боли.
|
|
|
Суммируя экспериментальные данные и результаты клинических наблюдений, можно заключить, что ноцицептивные стимулы различной природы вызывают, как правило, гипертензивные и тахикардические реакции, сопровождающиеся разнонаправленными изменениями органного кровотока и биохимическими проявлениями симпатико-адреналовой активации [Розонов Ю.Б., 1976; Зайцев А.А. и др., 1980; Дмитриев А.В., 1982; Игнатов Ю.Д., Зайцев А.А., 1987; Kissin I. et al., 1983; Kuchner M., Alavi A., 1985; Freye E., 1987, и мн. др.]. Описаны также депрессорные ноцицептивные реакции, однако они возникали лишь на фоне сверхглубокого наркоза или под влиянием субстанций типа брадикинина, обладающих прямым сосудорасширяющим свойством [Фолков Б., Нил Э., 1976; Панов А.В., 1988]. У бодрствующих животных в ряде исследований выявлена четкая корреляция между сдвигами гемодинамики и эмоционально-аффективными проявлениями боли различного генеза [Медведев О.С., Степочкина Н.А.» 1977; Дмитриев А.В., 1982; Зайцев А.А., Бершадский Б.Г., 1984].
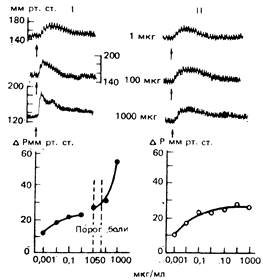
Конкретный нейрофизиологический субстрат реализации ноцицептивных вазомоторных рефлексов мало изучен. Предполагается, что ноцицептивная импульсация вызывает прессорные реакции, характерными особенностями которых являются крутой фронт нарастания и высокая амплитуда (рис. 12). Эти реакции возникают не в результате активации специализированных «болевых» рецепторов, а вследствие возбуждения тонких афферентов. Для развития ноцицептивных вазомоторных рефлексов необходимо, чтобы суммарный поток импульсов от данного рецептивного поля достиг некоторой критической интенсивности, которая определяется количеством возбуждаемых тонких волокон, частотой их разрядов и степенью удаленности данного поля от входа в спинной мозг [Хаютин В.М., 1980]. Показано, что селективное раздражение А-δ-афферентных волокон приводит к возникновению однокомпонентного симпатического разряда в почечном нерве и небольшим сдвигам артериального давления, а при активации С-волокон формируется вторая волна биоэлектрического ответа и резко увеличиваются амплитуда и скорость нарастания прессорной реакции артериального давления [Богданов Б.Г., 1986; Mense S., 1983]. Однако С-волокна не являются единственными проводниками информации, ответственной за возникновение ноцицептивных прессорных реакций, поскольку такие реакции, вызванные растяжением кишечника, сохраняются в полном объеме после введения капсаицина, вызывающего дегенерацию тонких афферентных волокон [Lembeck F., Skotitsch G, 1982].
|
|
|
|
Рис. 12. Рефлекторные реакции артериального давления у наркотизированной уретаном кошки, вызванные поочередным введением ацетилхолина (I) и мехолина (II) в сосуды перфузируемой тонкой кишки (по В.М. Хаютину, 1980). Приведены дозы препаратов.
В основе гипертензивных сдвигов, запускаемых ноцицептивным воздействием, лежит активация симпатических механизмов различных уровней ЦНС. Примечательно, что гемодинамические ноцицептивные реакции отчетливо коррелируют с С-компонентом сомато- и висцеро-симпатических рефлекторных ответов. Однако это вовсе не означает, что супрасегментарные вазомоторные механизмы играют основную роль в формировании ноцицептивных реакций артериального давления, поскольку в последние годы существенной ревизии подвергнуты представления о том, что С-ответы (С-симпатические рефлексы) интегрируются на супра-сегментарном, а А-рефлексы — на сегментарном уровне ЦНС. Установлено, что большинство компонентов симпатических рефлекторных ответов целиком или частично реализуются через спинной мозг [Хаютин В.М., 1980; Лебедев В.П., 1986].
|
|
|
Нейрофизиолргическим субстратом сегментарных гемодинамических реакций являются преганглионарные симпатические нейроны, локализованные преимущественно в боковом роге спинного мозга, а также в промежуточной зоне и вблизи центрального канала. Цитоархитектоника и свойства преганглионарных симпатических нейронов подробно изложены в ряде обобщающих работ [см.: Лебедев В.П., 1986; Janig W., 1982].
Аксоны этих нейронов не только выходят через передние корешки и достигают ганглиев, но и распространяются внутри спинного мозга, образуя проприоспинальный симпатический тракт. Посредством проприоспинальной системы преганглионарные нейроны активируются соматическими и висцеральными стимулами, и эта система участвует в генерализации симпатических рефлексов. Именно вовлечение через систему проприоспинальных связей преганглионарных нейронов различных сегментов определяет сдвиги системной гемодинамики, сопряженные с изменениями регионарного кровообращения, прежде всего в скелетных мышцах и мезентериальных сосудах, как наиболее массивных (емких) сосудистых зонах, которые необходимы для реализации поведенческих реакций избегания или устранения вредоносного стимула. Очевидно, что структура гемодинамической реакции при этом достаточно универсальна и вполне соответствует картине, сопровождающей типичную реакцию типа «страх—ярость», которая включает повышение артериального давления, увеличение сердечного выброса, сужение сосудов почек и желудочно-кишечного тракта с одновременным расширением сосудов скелетных мышц, связанным с активацией системы холинергической вазодилатации [Вальдман А.В. и др., 1979]. Можно предположить, что генерализация, и вместе с тем отчетливая регионарная дифференцировка сдвигов кровообращения при боли, обусловлена не только активацией проприоспинальной системы интермедио-медиального ядра, но и синергичным функционированием проприоспинальных систем спинного мозга разной модальности [Костюк П.Г., Преображенский Н.Н., 1975; Cervero F., 1983, 1985]. Об этом же свидетельствует прямая зависимость между выраженностью и генерализацией моторных реакций и реакций сердечно-сосудистой системы при увеличении интенсивности ноцицептивного раздражения.
|
|
|
Несомненно, что сам сегментарный уровень вазомоторной регуляции не в состоянии полноценно регулировать кровообращение в норме и тем более при боли. Изменение кровообращения в разных регионах организма в рамках генерализованной симпатико-адреналовой активации при боли требует тонких модулирующих влияний, реализуемых структурами супрасегментарного и прежде всего — бульбарного уровня. Многочисленными исследованиями доказано существование специфических «вазомоторных» нейронов в ядрах продолговатого мозга, получающих проекции от «собственных» рефлексогенных зон сердечно-сосудистой системы [подробнее см.: Фолков Б., Нил Э., 1976; Лебедев В.П., 1986]. Установлено существование специализированных нисходящих проекций бульбарных нейронов к симпатическим нейронам спинного мозга. Примечательно также, что восходящий поток ноцицептивных сигналов интегрируется как раз в структурах бульбарного отдела головного прессорных сдвигов артериального давления при боли и в значительной мере определяет индивидуальные особенности гемо динамического обеспечения нопицептивных реакций. Более того, имеющиеся в настоящее время данные позволяют рассматривать собственный гомеостатический механизм сердечно-сосудистой системы — барорецепторные рефлексы, — как универсальную систему «генерализованного торможения», при запуске которой, наряду с ослаблением гипердинамических сдвигов кровообращения, развивается угнетение психомоторных функций и аверсивных реакций различной, и в том числе ноцицентивной, природы [подробнее см.: Randich A., Maixner W., 1984]. Сердечно-легочные и синоаортальные бароафференты модулируют нейротранссмиссию в различных образованиях продолговатого мозга: гигантоклеточном ядре, ядре солитарного тракта, «синем пятне» и некоторых других. В этих же структурах представлены соматические и висцеральные входы от спиноталамического и спиноретнкулярного трактов, которые проецируются из I, IV, V и VII пластин заднего рога спинного мозга и осуществляют передачу ноцицептивных сигналов к гипоталамо-лимбическому комплексу и коре. Широкие анатомические связи различной модальности на бульбарном уровне являются основой установленного в электрофизиологических экспериментах барорефлекторного торможения болевой импульсации как от кожи, так и от внутренних органов. Очевидно, что ведущая роль в этих процессах принадлежит ядру солитарного тракта, что подтверждается высоким содержанием в нем опиатных рецепторов, а также развитием налоксонзависимой аналгезии при микроинъекциях опиатов в это ядро [Lewis J. et al., 1987].
|
|
|
Приведенные данные доказывают тесную взаимосвязь сдвигов кровообращения при боли с ее эмоционально-аффективными проявлениями. Вместе с тем они не позволяют ответить на вопрос об особенностях формирования разномодальных ноцицептивных реакций в рамках вегетомоторного сопряжения, имеющий стратегическое значение для оптимизации медикаментозной терапии болевых синдромов. Важно подчеркнуть, что уже сам факт развития гипертензии и тахикардии под влиянием ноцицептивных стимулов у наркотизированных животных, а также данные клинической анестезиологии о резистентности гемодинамических и эндокринных сдвигов к болеутоляющим средствам позволяют предполагать существование таких особенностей.
Определенную ясность в этот вопрос внесли исследования последних лет, посвященные роли антиноцицептивных систем головного мозга в регуляции кровообращения.
Весьма вероятно, что антиноцицептивные системы обладают тонической активностью, поскольку разрушение ЦСВ среднего мозга у крыс приводит к тахикардии [Bernet F., Denimal J., 1982]. У децеребрированных кошек прессорные ноцицептивные рефлексы развиваются только при сохранении связи продолговатого мозга со средним и когда от продолговатого мозга отделена его ростральная часть, примыкающая к мосту [Хаютин В.М., 1980]. Следовательно, часть антиноцицептивных систем, локализующаяся в области понтобульбарного соединения, способна тонически подавлять ноцицепгивные вазомоторные рефлексы без искусственной активации нисходящих путей. Активация антиноцицептивных структур вызывает не только аналгезию, но и изменения спонтанного поведения и фоновой гемодинамики, в частности артериальную гипертензию [Игнатов Ю.Д., Зайцев А.А., 1987; Janss A. et ai., 1987; Depauiis A. et al., 1988]. Электрическое раздражение околоводопроводного серого вещества среднего мозга, вызывающее стимуляционную аналгезию у бодрствующих кошек, может одновременно и зависимо от интенсивности центральной стимуляции уменьшать гемодинамические ноцицептивные реакции [Зайцев А.А. и др., 1982]. Такое же действие выявлено и при опосредованной активации антиноцицептивных зон головного мозга, которая происходит при акупунктурной аналгезии и при аналгезии, вызванной транскраниальной электростимуляцией [подробнее см.: Игнатов Ю.Д, Зайцев А.А., 1987; Лебедев В.И. и др., 1987]. В клинических условиях транскраниальная электроаналгезия предупреждает вегетативные и гормональные сдвиги при болезненных диагностических манипуляциях, сохраняющиеся на фоне медикаментозного анестезиологического пособия у различных групп больных, а у пациентов с лабильной артериальной гипертензией — вызывает отчетливое длительное снижение систолического и диастолического артериального давления. В экспериментах на бодрствующих кроликах транскраниальная аналгезия угнетала прессорные реакции артериального давления, вызванные раздражением А- и С-волокон седалищного нерва, уменьшала амплитуду позднего компонента соматосимпатического рефлекса, и в меньшей степени — подъем артериального давления в ответ на окклюзию сонных артерий.
Вместе с тем выраженность стимуляционной аналгезии зависит от интенсивности раздражения, и она пропорциональна повышению системного артериального давления [Игнатов Ю.Д., Зайцев А.А., 1987]. По нашим данным, аналгезия, аналогичная стимуляционной, возникала и при повышении артериального давления мезатоном, оказывающим сугубо периферическое адреномиметическое действие (рис. 13). В данном случае аналгезия причинно связана с гипертензией, поскольку проявлялась она только в период подъема артериального давления и прогрессировала при последовательном ступенчатом его повышении при повторном введении мезатона. Характерно, что налоксон не устранял аналгезию, вызванную мезатоновой гипертензией, и лишь частично ослаблял стимуляционную аналгезию, возникающую на фоне выраженной гипертензии.
Гипертензия, развивающаяся на фоне стимуляционной аналгезии, указывает на возможность синергичного функционирования собственно болевого (антиноцицептивные системы) и собственно сердечно-сосудистого (барорефлексы) гомеостатических механизмов. По имеющемся данным [подробнее см.: Randich A., Maixner W., 1984; Naranjo J., Fuentes J., 1985], при искусственной активации барорецепторов уменьшается выраженность реакции избегания крыс на стимуляцию тройничного комплекса. У людей по мере формирования эссенциальной гипертензии обнаруживается снижение реактивности к раздражению пульпы зуба, аналогичные изменения характерны для животных с различными формами гипертензии, причем билатеральная синоаортальная денервация снижает у них болевые пороги. Установлено, что структуры бульбарного уровня, такие как парагигантоклеточное ядро или гигантоклеточное, получающее ноцицептивные входы и непосредственно участвующие в реализации барорефлекторных влияний [Lewis J. et al., 1987; Lovick Т., 1987], имеют прямые проекции как от нейронов ядра солитарного тракта, так и от нейронов большого ядра шва. Именно эти конвергентные бульбарные образования участвуют в формировании опиатной и неопиатной аналгезии и угнетении функции сердечно-сосудистой системы.
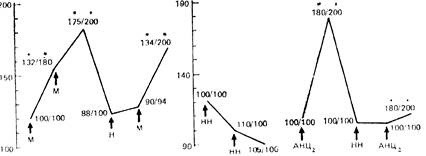
Рис 13. Изменения ноцицептивных реакций в зависимости от сдвигов артериального давления, вызванных вазоактивными препаратами.
Числа у графиков — латентный период отдергивания хвоста (в числителе) и порог вокализации (в знаменателе) и % от контроля; по оси ординат - уровень артериального давления (мм рт ст.) Стрелками указано введение мезатона (М), натрия нитропруссида (НН) налоксона (Н — 1 мг/кг). АНЦ; — раздражение среднего мозга с интенсивностью. вызывающей максимальную стимуляционную аналгезию. * — р<0.05 по сравнению с контролем.
Существует несомненная связь между барорефлекторными механизмами регуляции болевой чувствительности и процессами, возникающими при активации антиноцицептивных зон головного мозга. Очевидно, что при их градуально усиливающемся раздражении последовательно запускаются механизмы, кардинально различающиеся по нейрохимическому профилю. Начальные проявления стимуляционной аналгезии формируются за счет опиоидергических систем нисходящего торможения, в то время как усиление аналгезии опосредуется неоплатными механизмами, одновременно обеспечивающими гемодинамические предпосылки для избегания аверсивного раздражения. Развивающаяся при этом артериальная гипертензия может быть фактором, запускающим через барорецепторы высокого давления барорефлекторные механизмы регуляции эмоционально-аффективных проявлений боли. С другой стороны, равновероятна возможность непосредственной активации центральных звеньев барорефлексов в процессе раздражения антиноцицептивных систем головного мозга. Это положение доказывается нашими данными, показывающими, что стимуляционная аналгезия сохранялась в полном объеме на фоне введения натрия нитропруссида в дозе, не изменяющей фонового артериального давления, но препятствующей его повышению при раздражении антиноцицептивных зон мозга, т.е. натрия нитропруссид исключал активацию барорефлексов через периферические сосудистые рецепторы (см. рис. 13). Следовательно, эти данные позволяют предполагать взаимодействие барорефлексов и антнноцицептивных систем на супрасегментарном уровне ЦНС.
Важно подчеркнуть, что болеутоляющая функция барорецепторных рефлексов проявляется лишь в достаточно узком диапазоне и по мере усиления ноцицептивного воздействия происходит как бы ускользание болевых импульсов от бароконтроля, их «прорыв» к вышележащим структурам головного мозга, интегрирующим вокализацию и другие типичные компоненты генерализованной эмоционально-аффективной реакции на боль. Учитывая структурную иерархию и последовательность включения при боли разных гомеостатических механизмов, можно предположить, что «анальгетические системы» как раз определяют диапазон барорефлекторной регуляции, ограничивая ее эффективность при очень сильных воздействиях. Подобный характер взаимоотношений двух гомеостатических систем оправдан с точки зрения биологической целесообразности, поскольку устранение бароконтроля является одним из ведущих механизмов формирования гипердинамических сдвигов кровообращения при боли, необходимых для устранения аверсивного воздействия.
Представленный фактический материал свидетельствует о сложности и многочисленных особенностях нейрофизиологических механизмов формирования и регуляции сердечно-сосудистых реакций при боли. Главная из них заключается в самой функциональной значимости ноцицептивной информации, приводящей различные системы организма в состояние максимальной готовности и, следовательно, создающей крайне сложную взаимосвязь вазомоторных механизмов — нейрогенных и гуморальных. Не до конца понятно и, по-видимому, необычно участие барорефлекторных механизмов в регуляции комплексной ноцицептивной реакции организма. Из имеющихся данных следует, что барорефлексы регулируют не только гемодинамические, но и поведенческие ноцицептивные реакции, причем их «антиноцицептивное» действие проявляется как при естественной активации барорецепторов при повышении артериального давления, так и при первичном запуске антиноцицептивных систем среднего мозга.
Ключевую роль в регуляции различных проявлений боли играют антиноцицептивные системы головного мозга, которые, с одной стороны, тормозят негативные эмоции и их моторные компоненты, а с другой — мобилизуют вегетативные функции на избегание повреждающего воздействия. На наш взгляд, тесная взаимосвязь между стимуляционной аналгезией и артериальным давлением отражает разобщенность механизмов регуляции разномодальных функций при боли как крайнем варианте аверсивного ощущения. Недаром отмечают сходство сдвигов гемодинамики (повышение артериального давления, тахикардия, усиление мышечного кровотока) при электрической или химической (гомоцистеиновая кислота) активации среднего мозга с изменениями кровообращения при типичной «защитной реакции» [Duggan А., 1982; McDongall A. et а1., 1985]. По крайней мере, это оправдано с точки зрения биологической целесообразности и позволяет рассматривать антиноципептивные системы не только как механизмы подавления боли, но и как механизмы, обеспечивающие предпосылки к ее активному избеганию с целью устранения повреждающего воздействия.
Таким образом, несмотря на тесную взаимосвязь механизмов формирования эмоциональных, моторных и гемодинамических проявлений боли не вызывают сомнения особенности реализации ее разномодальных компонентов, определяемые на уровне нейромедиаторных и даже рецепторных механизмов. Полинейрохимизм процессов формирования реакций сердечно-сосудистой системы при боли изучен в значительно меньшей степени, чем нейромедиаторные механизмы регуляции эмоциональных и моторных ноцицептивных реакций. Несмотря на сформировавшееся мнение о важной роли в регуляции кровообращения в норме и при патологических состояниях серотонинергических и ГАМКергических систем [подробнее см.: Шмуйлович Л.М., Кудрин А.Н., 1987; De Feudis F., 1983; Krstic M„ 1985; Syrnoens J., Janssens M., 1986; Bom A. et al., 1988], практически отсутствуют данные, позволяющие обосновать их значение в контроле гемодинамики при боли. Подробнее исследованы опиоид- и адренергические механизмы, что, по-видимому, обусловлено их базовой ролью в регуляции болевой чувствительности и сердечно-сосудистых функций соответственно.
В ряде обзоров последних лет обсуждается значение опиатных нейропептидов в вазомоторных механизмах головного и спинного мозга, в функционировании нисходящих систем ствола головного мозга, барорефлексах и других процессах, определяющих вегетомоторное сопряжение [Вальдман А.В., Медведев О.С., 1982; Romagnano M, Hamill R., 1984; Howe P., 1985; Laubie M., Schrnitt H., 1985; Campbell G., 1986]. Радиоиммунологически показано совместное распределение на всем протяжении спинного мозга млекопитающих симпатических нейронов и энкефалинсодержащих волокон, что рассматривается как морфологическая основа опиоидергической регуляции вегетативных функций. Энкефалины, эндорфины и опиатные рецепторы выявлены также в других структурах ЦНС, участвующих в контроле за кровообращением. Предполагается, что метэнкефалин определяет «прессорные» системы организма, тогда как β-эндорфин составляет основу «депрессорных» систем.
По-видимому, опиоидергические системы регуляции гемодинамики обладают незначительной тонической активностью, поскольку, по данным большинства авторов, налоксон в дозах, блокирующих различные типы опиатных рецепторов, не вызывает закономерных сдвигов артериального давления и частоты сердечных сокращений у нормотензивных животных и людей [Вальдман А.В., Медведев О.С., 1982; Fuehmayor N.. Cubeddu L., 1986]. Это представление подтверждается результатами многочисленных экспериментов и клиническими наблюдениями, в которых на фоне морфина, фентанила и других опиоидов в оптимальных анальгетических дозах наблюдается стабильность или незначительная депрессия фоновых показателей системной и регионарной гемодинамики [Ghoneim M et а1 1984; Goldstein G., 1985; Freye F, 1987].
Использование налоксона позволило установить важную роль опиоидергических механизмов в колебаниях артериального давления в процессе физиологического сна и при эмоциональном стрессе, в формировании артериальных гипертензий, гипотензии при травматическом, геморрагическом, септическом и других видах шока [Bell J. et а1., 1985; Мееr К. et а1., 1986; Naranjo J. et а1., 1986; Ludbrook J., Rutter P., 1987]. У наркотизированных собак налоксон не изменяет гипертензивные сдвиги при стимуляции n. saphenus, но в условиях ваготомии и денервации синусов усиливает прессорные реакции артериального давления, вызванные раздражением афферентных волокон группы А-б. У интактных и спинальных животных налоксон усиливает вокализацию и повышает амплитуду С-рефлекса при стимуляции соматического нерва. Следовательно, не вызывает сомнений участие опиоидергических механизмов в формировании сдвигов системной гемодинамики при аверсивных воздействиях. Однако до настоящего времени противоречивы представления не только об их нейрофизиологической организации, но и о функциональной направленности. Предполагается, что гемодинамические эффекты опиатов реализуются преимущественно через супрасегментарные структуры, поскольку децеребрация не изменяет, а спинализация ослабляет блокирующее влияние налоксона на гипотензивное и брадикардическое действие морфина [Feldberg W., 1980]. Существенное значение придается и опиоидергическим процессам в спинном мозге [Marshally D., Buccafusco J., 1985; Maurette P. et al„ 1985].
Следует подчеркнуть, что гемодинамические ноцицептивные реакции справедливо рассматриваются в качестве коррелятов эмоционально-поведенческих проявлений боли, обеспечивающих нутритивную функцию и, следовательно, определяющихся во многом активностью скелетной мускулатуры. С этих позиций сформировалось аксиоматическое представление о принципиальной идентичности опиоидергических механизмов регуляции разномодальных компонентов болевых реакций. Анализ собственных результатов (табл. 1) и данных литературы показывает, что на фоне торможения поведенческих проявлений боли гемодинамические сдвиги либо достоверно не изменяются, либо даже могут увеличиваться, причем резистентность сердечно-сосудистых ноцицептивных реакций характерна для всех основных болеутоляющих средств (морфина, промедола, фентанила, пентазоцина и др.) в диапазоне анальгетических доз (подробнее см. гл. 2). Эта диссоциация не зависит от природы болевых стимулов — механических, электрических, термических, химических. Приведенные результаты позволяют предполагать, что процессы опиоидергической регуляции разномодальных функций организма могут быть разобщены и что именно нейрохимическая разобщенность определяет особенности нейрофизиологических процессов регуляции гемодинамики при боли [Игнатов Ю.Д., Зайцев А.А., 1984; Игнатов Ю.Д. и др., 1987].
Таблица 1
Болеутоляющее действие опиатов и апиоидов и их влияние на ноцицептивные сдвиги артериального давления
| Вещество, доза, способ введения | Показатели в % от контроля | ||||
| Латентный период от-дергивания хвоста | Порог вокализации | Амплитуда прессорных реакций АД | Уровень АД | ||
| Внутрибрюшиино (мг/кг): | |||||
| Морфин | 130* | 175* | 111* | 93* | |
| 175* | 210* | 120* | |||
| промедол | 215* | 275* | 148* | ||
| фентанил | 0.03 | 165* | 190* | ||
| Пентазоцин | 195* | 126* | |||
| бупренорфин | 0,025 | 185* | 335* | 46* | |
| Трамадол | 26—50 | 170* | 355* | 117* | 109* |
| В желудочек головного мозга (мкг): | |||||
| морфин | 180* | 285* | 95* | ||
| ДАГО | 0,5 | 240* | 200* | 131* | |
| ДАДЛ | 180* | 205* | 133* | ||
| бремазоцин | 10—30 | 125* | |||
| фенциклидин | 20—50 | 125* | |||
| Интратекально (мкг): | |||||
| морфин | 175* | 129* | |||
| ДАГО | 0,5 | 170* | 160* | 69* | |
| ДАДЛ | 245* | 130* | 133* | ||
| Бремазоцин | 10—30 | 165* | 131* | ||
| фенциклидин | 20—50 | 195* | 119* |
*р<0,05.
Примечание. Цифры отражают эффекты соединений через 30 мин после внутрибрюшинного введения (фентанил — через 15 мин после внутривенного введения) и через 5 — 15 мин. после микроинъекции.
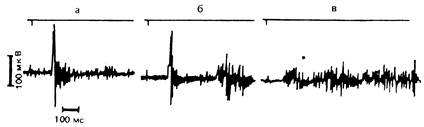
Рис. 14. Влияние морфина при интратекальном введении ненаркотизированным кошкам на рефлекторные ответы в почечном нерве при одиночном раздражении малоберцового нерва. а — контроль; б, в — морфин в дозах 5 и 100 мкг соответственно.
Наши эксперименты с локальными микроинъекциями анальгетиков в боковой желудочек головного мозга и под оболочки спинного мозга позволили выявить уровни ЦНС, на которых опиоидергическая регуляция разномодальных проявлений боли имеет неодинаковую направленность. Как видно из рис. 14, только при интратекальном введении морфин уже в субанальгетических дозах избирательно увеличивал С-компонент соматосимпатического рефлекса, а в анальгетических дозах вызывал в почечном нерве высокочастотный разряд, не разделяющийся на коротко- и длительнолатентные компоненты. Характерно, что в этих дозах морфин не уменьшал гемодинамические и биоэлектрические сдвиги, вызванные ритмическим ноцицептивным раздражением афферентных нервов, но индуцировал выраженную гипертензию и усиливал фоновую симпатическую активность в почечном нерве. Полученные данные позволили предположить, что такое симпатоактивирующее действие морфина обусловлено его влиянием непосредственно на симпатические вазомоторные механизмы сегментарного уровня. И действительно, у спинальных животных морфин вызывал аналогичные изменения гемодинамических показателей и биоэлектрической симпатической активности и, более того, увеличивал их сдвиги при ноцицептивном раздражении чревного нерва (рис. 15).
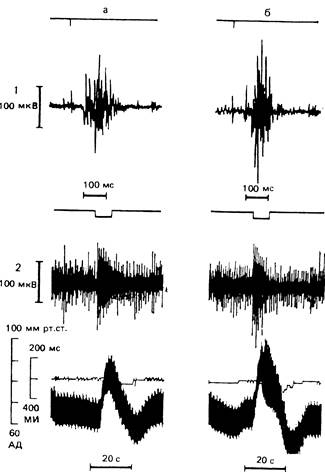
Рис. 15. Влияние морфина при интратекальном введении спинальным кошкам на реакции артериального давления, межсистольных интервалов и изменения биоэлектрической активности в почечном нерве при ноцицептивном раздражении чревного нерва.
а — контроль; б — морфин в дозе 100 мкг, 1 (сверху вниз) — отметка одиночного раздражения чревного нерва, активность в почечном нерве, 2 (сверху вниз) — отметка ритмического раздражения чревного нерва, активность в почечном нерве, межсистольные интервалы (МИ). артериальное давление (АД).
Эти результаты дали основания считать, что симпатоактивирующий эффект, присущий морфину и морфиноподобным анальгетикам, является главной причиной резистентности ноцицептивных гемодинамических реакций к опиатам и опиоидам. Ведущее значение в его формировании принадлежит вазомоторным механизмам спинного мозга. Учитывая значительную временную дисперсию залпа С-афферентных волокон и обширное распространение импульсов по спинному мозгу вплоть до бульбарных структур [подробнее см.: Хаютин В.М. и др., 1977; Лебедев В.П., 1986], можно полагать, что наркотические анальгетики активируют в первую очередь систему генерализации симпатических рефлексов. В пользу такого предположения свидетельствуют результаты экспериментов на спинальных животных. В этих условиях, когда устранены нисходящие модулирующие влияния и снижены функциональные возможности проприоспинальной системы, отчетливо проявилась способность морфина усиливать ноцицептивные изменения артериального давления и симпатической активности.
Следует упомянуть, что в наших экспериментах, в которых осуществляли деструкцию ядер шва и нисходящих серотониновых терминалей посредством интрацистернального введения 5,7-дигидрокситриптамина [Игнатов Ю.Д. и др., 1988], выявлено уменьшение ноцицептивных реакций артериального давления. На этом фоне морфин оказывал «парадоксальное» действие, проявляющееся еще большим ослаблением гипертензивных сдвигов. По-видимому, именно серотонинергические механизмы могут быть ключевыми в реализации симпатоактивирующего действия морфина и морфиноподобных анальгетиков.
Для детального анализа рецепторной организации опиоидергических механизмов регуляции боли на разных уровнях ЦНС мы использовали селективные агонисты 4 основных типов опиатных рецепторов — μ- (ДАГО), 6 (ДАДЛ), х (бремазоцин) и б (фенциклидин). Сопоставляли их эффекты при внутрижелудочковом и интратекальном способах введения в тестах отдергивания хвоста и вокализации, и регистрировали ноцицептивные сдвиги артериального давления и частоты сердечных сокращений. Такой подход позволял анализировать корреляцию супра- и сегментарных механизмов формирования боли и гемодинамических ноцицептивных реакций и процессы их взаимодействия. Как видно из табл. 1, морфин, агонисты х- и а-рецепторов при внутрижелудочковом введении в анальгетических дозах не изменяли величину прессорных реакций артериального давления, а ДАГО и ДАДЛ даже достоверно их увеличивали. Известно, что микроинъекции ДАГО и другого μ- агониста морфицептина в боковые желудочки или структуры гипоталамуса сопровождаются длительным повышением артериального давления и кратковременной брадикардией, сменяющейся учащением сердцебиений [Pfeiffer A. et al., 1983; Widy-Tyszkiewicz К. et al., 1986]. Эти эффекты устранялись налоксоном и β-фуналтрексамином. Поскольку ДАГО при аналогичном способе введения улучшает восстановление артериального давления и стимулирует работу сердца у животных с массивной кровопотерей, полагают, что на супрасегментарном уровне μ-опиоидергичсские системы выполняют вегетоактивируюшую функцию. Следует подчеркнуть, что супрасегментарный уровень вазомоторной регуляции характеризуется сложным взаимодействием δ-опиоидергических механизмов и нейрохимических процессов иного профиля. Так, прессорный и гахикардичсскнй эффекты ДАДЛ после введения в желудочки головного мозга бодрствующим крысам не устраняются налоксоном [McDonald W. et al., 1984]. В настоящее время представляется дискуссионным сформировавшееся около 10 лет назад мнение об активирующем эффекте х-опиатных механизмов в регуляции кровообращения, поскольку использование высоко-селективных агонистов х-рецепторов дали противоречивые результаты. Вещество U 50.488 Н, микроинъецированное в ядро одиночного пучка, вызывало артериальную гипертензию и повышение уровня вазопрессина в плазме у крыс, а другой х-агонист MR 2034, введенный в гипоталамус крыс, не вызывал сдвигов артериального давления и частоты сердечных сокращений [Pfeiffer А. et al, 1983; Carter D., Lightman S, 1985].
Наши данные об опиатных агонистах свидетельствуют об отсутствии корреляции между их гемодинамическими эффектами и влиянием на супрасегментарныс механизмы регуляции боли. Из табл. 1 видно, что болеутоляющее действие морфина,ДАГО и ДАДЛ формируется не только на сегментарном и супрасегментарном уровнях, но и за счет усиления нисходящего торможения нейронов спинного мозга. Депримирующее влияние ДАГО и ДЛДЛ на реакцию вокализации сопровождается усилением гемодинамических ноцицептивных реакций, в то время как морфин не изменяет их выраженности. Бремазоцин и фенциклидин, которые в отличие от морфина не усиливают нисходящее торможение, также не изменяют прессорные реакции артериального давления. По-видимому, супрасегментарные опиоидергические системы различного рецепторного профиля не играют важной роли в регуляции артериального давления при боли.
При интратекальном введении опиатных агонистов только ДАГО одновременно с аналгезией вызывал торможение ноцицептивных сдвигов артериального давления, а мор<
|
|
|
