 |


Глава 6 Биохимия микроорганизмов
|
|
|
|
Возможности микробов осуществлять химические реакции почти безграничны. Они способны создавать органические вещества из простых элементов — в этом проявляется их синтезирующая активность; могут также расщеплять самые сложные соединения на простые элементы и минеральные соли. Микробы осуществляют различные функции, однако самая важная на Земле - это их участие в круговороте веществ.
6.1 Круговорот веществ в природе
Участие микробов в круговороте веществ в природе— необходимое условие существования жизни на Земле. Обеспечивая минерализацию углерода из органических соединений растений, микробы поддерживают равновесие углекислого газа. Почвенным бактериям и грибам принадлежит огромная роль в регенерации СО2. В процессе минерализации органических форм углерода микробы почвы и воды не только переводят его обратно в СО2, но возвращают в круговорот веществ и остальные биоэлементы — азот, фосфор и др., которые лимитируют рост растений. Микробы участвуют в круговороте Р, N, S и других элементов не менее интенсивно, чем в круговороте углерода.
Зеленые растения, фиксируя СО2 при фотосинтезе, образуют прежде всего сахара и родственные им соединения. Основная масса их временно отлагается в виде полимеров. Из-за преобладания полисахаридов среди продуктов ассимиляции у зеленых растений сахара играют очень большую роль в качестве питательных веществ для всех организмов, зависящих от органических источников питания.
ПРЕВРАЩЕНИЕ ОРГАНИЧЕСКИХ ВЕЩЕСТВ, НЕ СОДЕРЖАЩИХ АЗОТ
Углерод в природе претерпевает сложные превращения в виде непрерывных процессов распада и синтеза, в которых принимают участие животные, растения и микроорганизмы.
|
|
|
Процессы брожения
К числу важнейших биохимических процессов, вызываемых микроорганизмами, относятся различные виды брожения — спиртовое, молочнокислое и др.
Брожением называют анаэробное разложение углеводов на продукты, которые далее не разлагаются без участия молекулярного кислорода. У различных организмов продукты брожения различны и зависят в основном от набора ферментов и от внешних условий.
Спиртовое брожение. Необходимую для жизнедеятельности энергию дрожжи получают, расщепляя углеводы на спирт и СО2, т. е в процессе спиртового брожения. Спиртовое брожение протекает в анаэробных условиях или при ограниченном доступе воздуха. В присутствии же кислорода дрожжи ведут себя как аэробные организмы и необходимую энергию получают за счет дыхания, окисляя энергетический материал до углекислого газа и воды. Поскольку этот путь энергетически более выгоден, то в аэробных условиях дрожжи хорошо размножаются. Поэтому при производстве пекарских или кормовых дрожжей применяют аэрацию, т. е. продувание среды воздухом.
Спиртовое брожение, лежащее в основе технологии получения спирта, пива, вина, кваса и других продуктов, выражается в общем виде уравнением Гей-Люс-сака:
С6Н1206 -> СН3СН2ОН + 2СО2.
Теоретически из 180 г глюкозы можно получить 92 г этилового спирта и 88 г углекислого газа. В действительности наряду с ними образуются побочные продукты— глицерин, янтарная кислота и др. Процесс превращения сахара в спирт и углекислый газ сопровождается выделением тепла, т. е. является экзотермическим. На 1 грамм-молекулу сброженного сахара выделяется тепла 117,3-106Дж.

Превращение сахара в спирт в результате жизнедеятельности дрожжей является сложным ферментативным процессом, осуществляемым через ряд промежуточных стадий, каждая из которых катализируется особым ферментом. Большую роль в процессе спиртового брожения играет фосфорная кислота, обеспечивающая механизм переноса энергии.
|
|
|
Химизм спиртового брожения изучали русские уче
ные Л. А. Иванов, С. П. Костычев и А. Н. Лебедев и за
рубежные ученые Нейберг, Мейергоф, Гарден, Эмбден
и др.
На первом этапе процесса моносахариды (глюкоза) подвергаются фосфорилированию. При этом к молекуле сахара присоединяются два остатка фосфорной кислоты, которая в виде аденозинтрифосфорной кислоты (АТФ) содержится в клетках дрожжей. В результате реакции образуется сначала глюкопиранозо-6-фосфат, а затем фруктофуранозо-1,6-дифосфат, и глюкоза приобретает строение фруктозы. При этом АТФ превращается в АДФ.
Из схемы видно, что глюкоза превращается в фруктозу и остатки фосфорной кислоты (обозначенные на схеме буквой Р) присоединяются к фруктозе на концах молекулы. Образовавшийся фруктофуранозо-1,6-ди-фосфат под влиянием фермента альдолазы, содержащейся в дрожжах, распадается на две молекулы •— 3-фосфоглицериновый альдегид и. фосфодиокси-ацетон:
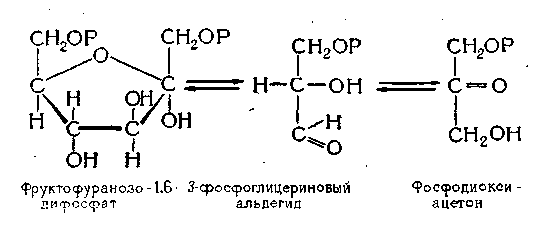
Эти фосфотриозы (каждая содержит по три углеродных атома) под влиянием фермента триозофосфатизо-меразы легко переходят одна в другую.
Дальнейшим превращениям подвергается 3-фосфоглицериновый альдегид. К нему присоединяется еще один остаток фосфорной кислоты и образуется дифос-фоглицериновый альдегид, который затем под действием фермента дегидрогеназы окисляется в 1,3-дифосфогли-цериновую кислоту. Активной группой этого фермента у дрожжей является НАД.

По мере использования 3-фосфоглицеринового альдегида — фосфодиоксиацетон образует новые количества 3-фосфоглицеринового альдегида.
1,3-Дифосфоглицериновая кислота вступает в реакцию с АДФ (образовавшейся на первой стадии процесса), отдает ей один остаток фосфорной кислоты и превращается в 3-фосфоглицериновую кислоту. Одновременно АДФ превращается в АТФ. Эту реакцию катализирует фермент фосфаттрансфераза. Под действием фермента фосфоглицеромутазы 3-фосфоглицериновая кислота далее превращается в 2-фосфоглицериновую кислоту. 2-Фосфоглицериновая кислота под действием фермента фосфопируватгидратазы (енолазы) отдает молекулу воды и превращается в фосфоенолпировиноградную кислоту. Это приводит к накоплению энергии в остатке фосфорной кислоты. Фосфоенолпировиноградная кислота отдает остаток фосфорной кислоты молекуле АДФ, благодаря чему образуется АТФ и енолпировиноградная кислота. Последняя под действием фермента фосфо-трансферазы превращается в более устойчивую кето-форму—пировиноградную кислоту.
|
|
|

При спиртовом брожении пировиноградная кислота расщепляется на ССЬ и уксусный альдегид под действием фермента пируватдекарбоксилазы:
СНзСОСООН - СО2+СН3СНО
Пировиноградиая Уксусный альдагид
Уксусный альдегид вступает во взаимодействие с НАД-Н2. В результате образуется этиловый спирт и регенерируется молекула НАД. Восстановление уксусного альдегида катализирует фермент алкогольдегидрогена-за, коферментом которого является НАД:

Образованием этилового спирта заканчивается цикл спиртового брожения.
Молочнокислое брожение - разложение сахара под действием молочнокислых бактерий - происходит с образованием молочной кислоты. Брожение открыто и впервые описано Пастером. Процесс протекает по уравнению
С6Н,2О6 -> 2СН3СНОНСООН.
Молочная кислота
Наряду с молочной кислотой образуются также побочные продукты. Это брожение чаще всего возникает в молоке, причем расщеплению подвергается молочный сахар - лактоза.
По характеру брожения различают две группы молочнокислых бактерий. Одни из них - гомофермен-тативные - образуют молочную кислоту почти как единственный продукт брожения. Другие же - гете-роферментативные наряду с молочной кислотой образуют значительные количества других продуктов: уксусную кислоту, этиловый спирт, углекислый газ, водород и некоторые ароматические вещества.
При молочнокислом брожении превращение сахара протекает так же, как при спиртовом брожении, до образования пировиноградной кислоты. Однако далее химизм эти/ двух типов брожения расходится. Из-за отсутствия у гомоферментативных молочнокислых бактерий фермента пируватдекарбоксилазы пировиноградная кислота не распадается на ацетальдегид и углекислый газ, а восстанавливается в молочную кислоту по уравнению
|
|
|
СНзСОСООН + НАД-Н2 ± СНзСНОНСООН + НАД.
Пировиноградная Молочная кислота
кислота
Эту реакцию катализирует фермент лактикодегидро-геназа и кофермент НАД. Как и при спиртовом брожении, в процессе окисления дифосфоглицеринового альдегида НАД восстанавливается и превращается в НАД-Н2. Передавая свой водород пировиноградной кислоте, она снова окисляется в НАД, а пировиноградная кислота восстанавливается в молочную кислоту.
Процесс гетероферментативного молочнокислого брожения изучен недостаточно. Бактерии, вызывающие это брожение, обладают ферментом пируватдекарбоксилазой, который вызывает распад пировиноградной кислоты на ацетальдегид и СО2. В качестве продуктов образуются молочная кислота, этиловый спирт, может быть и уксусная кислота.
Наиболее важное техническое значение имеют следующие молочнокислые бактерии:
сбраживающие лактозу и размножающиеся в молоке - молочнокислый и сливочный стрептококки. Они применяются для производства простокваши, творога, сметаны, масла и др.;
термофильные молочнокислые бактерии - используются при изготовлении кисломолочных продуктов, диетических продуктов и сыров;
не сбраживающая лактозу палочка Дельбрюка - применяется для производства молочной кислоты и при изготовлении жидких дрожжей.
В силосовании кормов и квашении овощей активное участие принимает огуречная палочка.
Пропионовокислое брожение заключается в превращении сахара, молочной кислоты или ее солей в пропионовую и уксусную кислоты по уравнению:
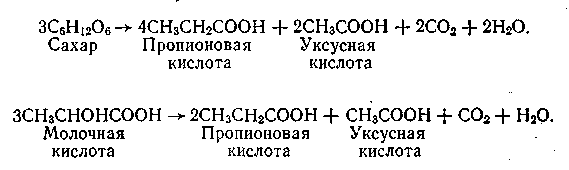
Реакции сопровождаются выделением энергии. По химизму Пропионовокислое брожение сходно со спиртовым и молочнокислым, и молочная кислота является как бы промежуточным продуктом. Вызывают это брожение пропионовокислые бактерии - короткие, бесспоровые, неподвижные, грамположительные палочки - анаэробы. Оптимальная температура развития их 30-35° С.
Маслянокислое брожение. Это сложный процесс превращения сахара бактериями. Он протекает в анаэробных условиях с образованием масляной кислоты, углекислого газа и водорода.
С6Н12О6 -> СН3СН2СООН + 2СО2 + 2Н2 + энергия. Сахар Масляная кислота
В качестве побочных продуктов маслянокислого брожения получаются бутиловый спирт, ацетон, этиловый спирт и уксусная кислота. Это брожение также было открыто Пастером в 1861 г.
Первые стадии маслянокислого брожения протекают так же, как спиртовое брожение. Происходит фосфори-лирование гексозы и распад ее на две молекулы фосфо-триоз, которые образуют затем ацетальдегид и COj.
|
|
|
В отличие от спиртового брожения, ацетальдегид далее конденсируется. При этом образуется соединение с четырьмя углеродными атомами:
СНзСНО + СНзСНО -> СН3СНОНСН2СНО.
Ацетальдегид Ацетальдоль
Образующийся ацетальдоль в дальнейшем превращается в масляную кислоту:
СН3СНОНСН2СНО -> СН3СН2СН2СООН.
Ацетальдоль Масляная кислота
Маслянокислые бактерии являются строгими анаэробами и могут размножаться при полном отсутствии или при очень малом количестве кислорода. Оптимальная температура развития бактерий 30-40° С. Эти бактерии широко распространены в природе, их постоянным местообитанием является почва, отложения озер, прудов, болот, остатки растений, навоз и др.
Ацетоно-бутиловое брожение. Химизм этого брожения сходен с маслянокислым. В результате брожения образуются бутиловый спирт, ацетон, этиловый спирт, масляная и уксусная кислоты и выделяются газы - СО2 и Н2.
Возбудителями брожения являются подвижные палочки клостридиум и ацетобутиликум. Это - спорообразующие анаэробы, способные сбраживать углеводы и крахмал. Так как ацетон и бутанол широко применяются в различных отраслях народного хозяйства, ацетонобутиловые бактерии используются для их получения в промышленных масштабах.
Ацетоно-этиловое брожение. Углеводы разлагаются с образованием ацетона, этилового спирта, углекислого газа и водорода. В виде побочных продуктов ацетоно-этилового брожения образуется небольшое количество масляной, уксусной и муравьиной кислот.
Возбудители ацетоно-этилового брожения принадлежат к факультативным анаэробным бактериям, образующим споры. Длина палочек достигает 4—10 мкм. Оптимальная температура развития 40-42° С, оптимальная реакция среды для размножения равна рН 8-9, для брожения рН 6-8. Эти бактерии способны гидролизовать пектиновые вещества и сбраживать различные углеводы.
Брожение клетчатки. Этот вид брожения также широко распространен в природе и заключается в разрушении клетчатки в анаэробных условиях с образованием масляной и уксусной кислот, а также газов: СО2, Н2 или СН4. Клетчатка (или целлюлоза) является очень сложным полисахаридом, химически очень устойчива. Под влиянием фермента целлюлазы клетчатка распадается на более простые соединения типа целлобиозы. В присутствии фермента целлобиазы целлюлоза переходит в глюкозу, которая сбраживается.
Целлюлаза и целлобиаза содержатся у некоторых микроорганизмов - грибов и бактерий.
Сущность брожения клетчатки раскрыта В. Л. Оме-лянским (1902), который выделил две разновидности целлюлозных бактерий: одни бактерии вызывают брожение клетчатки с образованием в основном водорода (водородное брожение), а другие - метана (метановое брожение). В первом случае образуются в равном количестве уксусная и масляная кислоты, а газообразных продуктов немного. Во втором случае образуется преимущественно уксусная кислота и много газообразных продуктов.
Бактерии Омелянского - длинные тонкие палочки, анаэробы, способные к спорообразованию. Клетки со спорами имеют плектридиальную форму (барабанные палочки). Оптимальная температура развития 30-35°С.
В почвах широко распространены термофильные целлюлозные бактерии, также вызывающие брожение клетчатки.
Брожение пектиновых веществ (близко к масляно-кислому брожению). Пектиновые вещества содержатся в значительном количестве в растениях и состоят из пектиновых кислот и углеводов. Гидролиз пектиновых веществ может происходить при нагревании в присутствии кислот и под действием специфических ферментов. Эти ферменты - протопектиназа, пектиназа и пектаза - содержатся в растениях и во многих микроорганизмах. Под действием этих ферментов пектиновые вещества расщепляются на составляющие компоненты, которые в свою очередь распадаются на более простые соединения: уксусную, масляную и муравьиную кислоты, этиловый спирт, СО2 и Н2.
Возбудителями пектинового брожения являются подвижные бактерии, относящиеся к факультативным анаэробам, образующим споры.
Пектиновое брожение в природе имеет огромное значение, так как результатом его является разрушение растительных остатков. Эти процессы постоянно происходят в почве и в воде.
Процессы окисления
Биохимические процессы, связанные с дыханием микроорганизмов, с использованием кислорода воздуха, относятся к окислительным процессам.
Во многих случаях конечными продуктами являются не только углекислый газ и вода, но и продукты неполного или частичного окисления органических веществ. Так, при окислении сахара получается лимонная и другие органические кислоты, при окислении этилового спирта - уксусная кислота и т. д. Аэробные окислительные процессы, вызываемые микроорганизмами, сложнее и разнообразнее, чем анаэробные, и многие из них до настоящего времени недостаточно изучены.
Продукты неполного окисления, образуемые одними микроорганизмами, в природных условиях используются другими микроорганизмами. Так, группы микроорганизмов, последовательно сменяющие одна другую, могут довести окисление органических веществ до- конечных продуктов - СО2 и воды. При этом органическое вещество подвергается полной минерализации.
Окислительным процессам в природе подвержены все органические вещества, даже такие очень устойчивые в химическом отношении, как углеводороды, древесина, жиры, воски и парафины.
Окисление углеводородов. Углеводороды являются стойкими соединениями и могут окисляться лишь некоторыми микроорганизмами.
Метановые бактерии, например, окисляют метан по уравнению
СН4 + 202 -»- С02 + 2Н2О + 9 • 102 Дж.
Метан.служит для бактерий источником энергии, а также материалом для построения органических веществ клетки.
Некоторые виды микроорганизмов окисляют аналогичным образом углеводороды жирного ряда - пропан, бутан, гексан и другие с образованием углекислого газа и воды:
. СН3СН3 + 3-уО2->2СО2 + ЗН2О + энергия.
Бактериальному окислению могут подвергаться также углеводороды жирного ряда с более длинной цепью углеродных атомов. Так, русский ученый В. О. Таусон выделил из почвы различные бактерии, использующие в качестве единственных источников углерода углеводороды нефти, бензина, керосина, различных парафинов и т. д. Эти вещества могут служить источником углерода для некоторых бактерий и грибов, и конечными продуктами окисления также являются углекислота и вода. Так как в качестве промежуточных продуктов обнаруживают спирты и органические кислоты, считают, что процесс окисления насыщенных углеводородов связан с дегидрогенизацией их и образованием ненасыщенных соединений.
Эти процессы вызываются аэробными микроорганизмами и связаны с потреблением кислорода. Более интенсивно они происходят при хорошей аэрации или при наличии в среде веществ, способных отдавать кислород для этих окислительных процессов. Наиболее доступны для окисления углеводороды, содержащие гидроксильные и карбоксильные группы.
Окисление жиров и высокомолекулярных кислот жир ного ряда Жирные кислоты, входящие в состав различных жиров, окисляются аналогичным образом. Попадая в почву и водоемы вместе с растительными и животными остатками, они разлагаются различными микроорганизмами: бактериями, актиномицетами и грибами. Наиболее энергично разлагают жиры некоторые пигментные бактерии - бактериум продигиозум (образуют красный пигмент), псевдомонас флюоресценс (зеленовато-желтый пигмент). Это - палочковидные, неспорооб-разующие бактерии, многие устойчивы к низким температурам. Микроорганизмы расщепляют жир на глицерин и жирные кислоты и окисляют их до углекислоты и воды. Расщепление жира начинается с его гидролиза при помощи фермента липазы на глицерин и жирные кислоты:
Ж + ЗН20 ->• С3Н5(ОН)з + ЗС18Нз602,
Жир Стеариновая кислота
Жирные кислоты постепенно окисляются до углекислоты и воды по схеме
С18Н3бО2 + 2602-> 18СО2 + 18Н2О + энергия.
Окисление этилового спирта в уксусную кислоту
Процесс впервые был описан в 1862 г. Пастером, который обнаружил на поверхности скисшего вина и уксуса палочковидные бактерии. Окисление этилового спирта в уксусную кислоту называют уксуснокислым брожением; оно протекает по следующему суммарному уравнению:
СН3СН2ОН + О2 -»- СНзСООН + Н2О + энергия. Этиловый спирт Уксусная кислота
Реакция окисления этилового спирта протекает в две стадии. Вначале образуется уксусный альдегид, который затем окисляется в уксусную кислоту:
2СН3СН2ОН + O2-v2CH3CHO + 2Н2О;
2СН3СНО -f Ог -> 2СН3СООН. 102
Уксуснокислое брожение вызывают уксуснокислые бактерии - палочковидные, бесспоровые, аэробные микроорганизмы.
В природе уксуснокислые бактерии широко распространены, встречаются на плодах, ягодах, в вине, пиве, квасе,и т. д. Они вызывают скисание вина и других напитков.
.Уксуснокислые бактерии могут также окислять другие спирты и альдегиды, например пропиловый и бутиловый спирты, с образованием соответствующих кислот - пропионовой и масляной.
Некоторые уксуснокислые бактерии окисляют сахара, имеющие альдегидную группу, до соответствующих кислот. Например, глюкоза может быть окислена уксуснокислыми бактериями до глюконовой кислоты:
2СН2ОН (СНОН) 4СНО + О2 ->- 2СН2ОН (СНОН) 4СООН
Глюкоза Глюконовая кислота
Это превращение глюкозы называется глюконо-вокислым брожением.
Некоторые уксуснокислые бактерии вызывают окисление многоатомных спиртов в кетоспирты или кетоса-хара.
Окисление шестиатомного спирта сорбита в сорбозу используется для синтеза аскорбиновой кислоты.
Таким же образом может осуществляться биохимическое окисление глицерина в диоксиацетон, представляющий собой ценный продукт:
2СН2ОНСНОНСН2ОН + 02 -»• 2СН2ОНСОСН2ОН + 2Н2О.
Глицерин Диоксиацетон
Уксуснокислое брожение вызывают уксусные бактерии, входящие в род ацетобактерий (Acetobacter). Отдельные виды отличаются один от другого подвижностью, размерами клеток, устойчивостью к спирту и др. Некоторые могут накапливать в среде до 9,5-11,5% уксусной кислоты. Оптимальная температура роста бактерий 20-35° С. Бактерии часто встречаются в виде длинных нитей. Многие образуют пленку - сплошную или островками, тонкую или толстую, хрящевидную.
Окисление углеводов плесневыми грибами с образованием органических кислот. Способностью окислять углеводы обладают многие плесневые грибы. При этом из моно- и дисахаридов, а также из других органических веществ образуются органические кислоты: уксусная, молочная, щавелевая, янтарная, фумаровая, яблочная, лимонная и др.
При окислении органических веществ плесневыми грибами часто образуется щавелевая кислота. Она может далее окисляться до СОг и воды. Мукоровые грибы образуют янтарную кислоту в качестве промежуточного продукта; из нее затем образуется фумаровая кислота.
Окисление микроорганизмами клетчатки и близких к ней соединений. В почве протекает важный процесс аэробного окисления клетчатки (целлюлозы), вызываемый бактериями, актиномицетами и грибами. Клетчатка сначала гидролизуется, а затем окисляется. Одновременно образуются высокомолекулярные органические кислоты и более простые оксикислоты.
При разложении клетчатки в качестве промежуточных продуктов образуются растворимые сахара и органические кислоты. Они служат питательными веществами для почвенных бактерий и способствуют повышению плодородия почвы.
Почвенные бактерии и грибы могут кроме клетчатки окислять пентозаны, пектиновые вещества и лигнин. Пентозаны содержатся в растительных тканях наряду с клетчаткой и легко окисляются многими микроорганизмами до СО2 и воды. Образующиеся при их гидролизе пентозы (сахара с пятью углеродными атомами) кроме аэробного окисления бактериями и грибами могут сбраживаться некоторыми видами молочнокислых бактерий с образованием молочной и уксусной кислот.
Лигнин представляет собой сложное и стойкое соединение и входит в состав одревесневшей ткани растений. Способностью разлагать его обладают лишь некоторые грибы и бактерии. Химизм разложения лигнина неизвестен.
ПРЕВРАЩЕНИЕ ОРГАНИЧЕСКИХ АЗОТСОДЕРЖАЩИХ СОЕДИНЕНИЙ
В состав белков каждого живого существа входит азот, являющийся жизненно необходимым элементом. Общее содержание азота в растительных и животных Организмах достигает 10—25 млрд. т. На синтез клеточного вещества растений ежегодно расходуется до 1,5 млрд, т азота. В атмосфере содержится еще больше азота, так как воздух на 4/s состоит из этого газа. До 80 тыс. т молекулярного азота находится в воздухе над каждым гектаром земли.
В почве запасы азота также очень велики. Тем не менее низкие урожаи растений происходят от недостатка азота. Причина заключается в том, что лишь небольшая часть азотсодержащих соединений доступна для питания растений. Для этой цели не годится не только молекулярный азот, но и органический, попадающий в почву в виде белковых соединений вместе с животными и растительными остатками. Сложные соединения должны быть превращены в более доступные формы, усвояемые растениями, или минерализоваться.
Аммонификацией называются процессы перехода сложных азотистых продуктов в соединения аммиака. Этим процессам принадлежит важнейшее место в круговороте азота в природе. Они осуществляются в результате жизнедеятельности различных бактерий, актиномицетов и плесневых грибов. При этом получаются различные конечные продукты.
Аммонификация мочевины происходит под действием особых бактерий, открытых Пастером в 1862 г.,— уробактерий. Они относятся к аэробам и хорошо развиваются только в среде с резко щелочной реакцией. Источниками углеродного питания служат соли различных кислот, моно- и дисахариды, декстрин и крахмал. В качестве источника азота они используют аммиачные соли или свободный аммиак. Дезаминирование мочевины происходит под влиянием фермента уреазы.
Уробактерии играют большую роль в разложении мочевины. Содержащегося в ней азота образуется на земном шаре более 20 млн. т в год, и он был бы недоступен растениям, если бы не деятельность уробактерий. Они представляют собой палочковидные и шаровидные бактерии. Палочковидные бактерии имеют длинные жгутики и образуют споры. Шаровидные уробактерии чаще объединены в пакеты по 4—8 клеток и также обладают длинными жгутиками.
Аммонифицирующие бактерии способны также минерализовать очень стойкое и сложное соединение - хитин, входящее в состав клеточных оболочек многих
микроорганизмов и панцирных покровов насекомых.
Аммонификация белковых веществ происходит под действием особых гнилостных бактерий, а также акти-номицетов и грибов. Белковые вещества попадают в почву в очень больших количествах вместе с остатками растений, животных и микроорганизмов. Разложение белка происходит во всех почвах и водоемах.
Так как большинство протеолитических ферментов микроорганизмов активны в щелочной среде, то в этих условиях гниение активируется. При низкой температуре гниение замедляется, а в условиях вечной мерзлоты вообще не происходит.
Микроорганизмы, вызывающие гниение, чрезвычайно широко распространены в природе. Их много в воде, воздухе и особенно в почве, где процессы гниения протекают очень интенсивно. В кишечнике человека и животных постоянно обитает ряд гнилостных бактерий. Энергичному протеканию в нем гнилостных процессов благоприятствует щелочная реакция среды.
В минерализации органических веществ почвы гниение имеет громадное значение. Оно играет санитарную роль, обусловливая биологическую очистку почвы, разрушение трупов и всевозможных органических отбросов.
Среди аэробных бактерий, участвующих в аммонификации белков, широко распространена почвенная бактерия бациллус микоидес (Bacillus mycoides) - небольшая спороносная палочка, перитрих. Колонии этой бактерии напоминают мицелий гриба.
Картофельный бацилл бациллус мезентерикус (Вас. mesentericus) является небольшой палочкой, имеющей жгутики, часто образует цепочки. Сенная палочка бациллус субтилис (Вас. subtitis) очень широко распространена в природе и вызывает энергичную аммонификацию белка. Является короткой подвижной палочкой, образует споры.
Среди факультативно-анаэробных бактерий, вызывающих аммонификацию белков, известна палочка протеус вульгарис (Proteus vulgaris). Бактерии эти подвижны, спор не образуют, легко меняют форму в зависимости от типа питательной среды. Бактерии очень широко распространены в природе. При разложении белка образуют сероводород и индол, углеводы сбраживают с выделением СО2, Н2 и органических кислот.
Кишечная палочка эшерихия коли (Escherichia co lt) также принимает активное участие в разложении белка, входит в состав микрофлоры кишечника человека и животных.
Среди анаэробных бактерий аммонификацию белка вызывают гнилостные бактерии бациллус путрификус (Bacillus putrificus) и др. При спорообразовании у них образуются характерные плектридиальные формы, или «барабанные палочки». При разложении белков дают большое количество газов.
В разложении белков кроме бактерий деятельное участие принимают и другие организмы - актиномицеты и различные грибы.
Конечный продукт гидролиза белка - аминокислоты - диффундируют внутрь микробных клеток и там подвергаются дезаминированию. Этот процесс приводит к отщеплению аммиака, в результате чего образуются органические кислоты и спирты по уравнениям:
R - СНМН2СООН + Н2О ->- R — СНОНСООН + NH3; R — CHNHjCOOH + '- 02 ч* RCOCOOH + NH3; R — CHNH2COOH + 2Н ->- R — СН2СООН + NH3;
При разложении белков образуются муравьиная, уксусная, пропионовая, масляная, валериановая и другие кислоты и высокомолекулярные спирты. В аэробных условиях кислоты полностью окисляются до СОг и Н2О; в анаэробных условиях полное окисление жирных кислот невозможно и они накапливаются в среде. Некоторые из них разлагаются на углекислый газ и метан.
Распад содержащихся в белке диаминокислот приводит к образованию ди ам инов, называемых также птомаинами, или трупным ядом. Он может вызывать отравление благодаря действию на организм ядовитых алкалоидов, входящих в его состав. Если в составе аминокислот содержится еще и сера (например, в цистине), то она при гниении выделяется в форме сероводорода или меркаптанов.
Большая часть получающихся при разложении белка продуктов в аэробных условиях подвергается дальнейшему окислению до СОа и Н2О. Такие продукты, как аммиак, сероводород и фосфорная кислота, могут использоваться микроорганизмами для построения белковых веществ своего тела.
Процесс гниения может развиваться также в пищевых продуктах и вызывать их порчу.
ПРЕВРАЩЕНИЕ МИНЕРАЛЬНЫХ ВЕЩЕСТВ
Минеральные азотсодержащие вещества в почве подвергаются воздействию большого числа микроорганизмов.
Нитрификация, или окисление аммиака, вызывается особыми нитрифицирующими бактериями и приводит к превращению его в азотистую и азотную кислоту. Интенсивная нитрификация происходит при аэробных условиях.
С. Н. Виноградский доказал, что процесс нитрификации является двухфазным и каждая фаза обусловливается деятельностью специфического микроба. В первой фазе происходит превращение аммиака в азотистую кислоту:
2NH3 + ЗО2 -> 2НМО2 + 2Н2О + 6,6- 1 02 Дж.
Этот процесс вызывают нитрозные бактерии. Вторая фаза нитрификации — окисление азотистой кислоты в азотную:
2.102 Дж.
Эта фаза нитрификации осуществляется группой нитратных бактерий. Нитрифицирующие бактерии являются типичными представителями автотрофных микроорганизмов. Они способны к ассимиляции углекислоты воздуха и необходимую им энергию получают в результате окисления аммиачных и азотистых солей. Даже небольшие количества таких органических соединений, как пептон, глюкоза и др., задерживают их развитие.
Нитрифицирующие микроорганизмы широко распространены в природе. В кислых почвах они почти не встречаются, в целинных почвах их мало и основное местообитание их - культурные почвы. Нитрификация энергичнее идет при широком притоке кислорода, оптимальной температуре 30-37° С и влажности 60-80% от полной влагоемкости почвы. В процессе нитрификации в почве образуются нитраты, которые являются лучшим источником азотистого питания растений.
Денитрификация. Часть нитратов в почве используется высшими растениями, часть вымывается водой, некоторое количество изменяется под влиянием микробов. Эти изменения заключаются в восстановлении их до нитритов и аммиака или даже до свободного азота.
Превращение нитратов в нитриты осуществляется многими плесневыми грибами из рода мукор, пенициллиум и неспороносными бактериями. Наличие в почве органических веществ, особенно углеводов, способствует восстановлению нитратов. Аэрация почвы, наоборот, задерживает денитрификацию.
Денитрифицирующие микроорганизмы окисляют углерод органических соединений, отнимая кислород от азотной кислоты, и получают при этом необходимую энергию.
Денитрифицирующие микроорганизмы широко распространены, они встречаются в почве, навозе, сточных водах. Главная масса этих бактерий в почве находится на глубине 10-15 см. Развитию их способствует щелочная реакция почвы, наличие влаги и ограниченный приток кислорода. Некоторые денитрифицирующие микроорганизмы могут выделять из нитратов до 30-90% газообразного азота.
Обеднение почвы связанным азотом и переход свободного азота в атмосферу уравновешиваются деятельностью другой группы микроорганизмов, которые фиксируют азот из воздуха и переводят его в связанном состоянии в почву.
Фиксация атмосферного азота. Фиксация атмосферного азота осуществляется в связи с деятельностью различных микроорганизмов и имеет огромное значение в природе и сельском хозяйстве. Известны два вида микроорганизмов, фиксирующих азот: клубеньковы е, развивающиеся на корнях бобовых растений, исвободноживущие.
Клубеньковые бактерии по морфологическим признакам делятся на две группы - перитрихиальную и моно-трихиальную. В культуре клубеньковые бактерии имеют вид палочек или кокков, иногда наблюдается вздутие или ветвление; в клубеньках форма клеток меняется. Клубеньковые бактерии - аэробные организмы, для питания используют углеводы растений и азот воздуха. Они широко распространены в почве, устойчивы к высыханию и солнечной радиации, играют большую роль в обогащении почвы азотом.
Атмосферный азот могут фиксировать также некоторые свободно живущие в почве бактерии и грибы. Представителями аэробных азотфиксирующих бактерий является азотобактер, анаэробных - клостридиум. Такие микроорганизмы широко распространены в почве. Активность их в значительной степени зависит от температуры и климатических условий. Низкая температура и высушивание почвы задерживают их развитие, известкование почвы и снабжение ее фосфатами способствуют фиксации азота.
Азотфиксирующие микроорганизмы играют огромную роль в сохранении и пополнении азотистых запасов почвы. На этом основано применение их как препаратов для повышения урожайности почв.
ПРЕВРАЩЕНИЯ СОЕДИНЕНИЙ СЕРЫ, ФОСФОРА И ЖЕЛЕЗА
Соединения серы, фосфора и железа, попадающие в почву вместе с растительными и животными остатками, претерпевают большие изменения.
Образование сероводорода. В почву попадает большое количество серы, и при разложении органических остатков она освобождается обычно в форме сероводорода. Источником серы являются белки, разложение которых приводит также к разрушению содержащих серу аминокислот — цистина, цистеина и метионина. Под влиянием гнилостных бактерий при этом образуется сероводород.
Сероводород образуется также при восстановлении солей серной, сернистой и серноватистой кислот, что вызывается особой группой десульфофицирующих (или сульфатредуцирующих) бактерий. К ним относятся спи-риллум десульфуриканс (Spirillum desulfuricans). Они
могут развиваться на средах, содержащих спирты и органические кислоты, и восстанавливать сульфаты.
Восстановление сульфатов может быть выражено суммарными уравнениями:
С6Н,2Об + 6Н2О -»- 6СО2 + 12Н2;
3H2S04 + 12Н2 -ч- 12Н2О + ЗН2;
С6Н,2О6 + 3H2SO4 -» 6С02 + 6Н2
