 |


Базовый комплекс упражнений 22 глава
|
|
|
|
| 
|
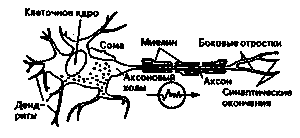
| Рис.1. Строение нервной клетки (нейрона). | Рис.2. Микрофотография клеточного тепа (сомы). |
 Рис. 3. Синаптические связи.
Рис. 3. Синаптические связи.
| 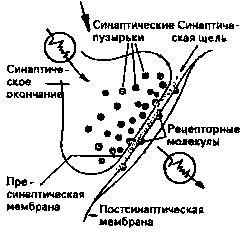 Рис.4. Синаптическая щель.
Рис.4. Синаптическая щель.
|
На рис. 3 мы видим многочисленные синаптические связи между синаптическими окончаниями и клеточным телом и дендритами соседней клетки.
Увеличенный фрагмент рис. 1 на рис. 4 показывает нам внутреннюю жизнь синаптического окончания, синаптическую щель, а также часть клеточной мембраны соседней клетки. Каждое синаптическое окончание наполнено маленькими пузырьками, каждый из которых содержит около 10 000 молекул химически активных веществ. которые называют нейротрансмиттерами. или передающим веществом.
Если рецепторная клетка возбуждается одним из ранее описанных раздражителей (или одна нервная клетка — другой), то определенное количество синаптических пузырьков перемещается в пресинаптической мембране и испускает свое содержимое в синаптическую щель. Возле постсинаптической мембраны соседней клетки это содержимое вступает в химическую реакцию с содержащимся в ней рецепторными молекулами, что приводит к деполяризации клеточной мембраны, которая расширяется за доли секунды вплоть до синаптических окончаний.
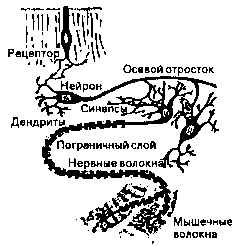
Рис.5. Синаптические связи.
При этом наступает непрерывное электрическое возбуждение нервной клетки: в состоянии покоя в ней присутствует отрицательный электрический потенциал приблизительно в -70 милливольт, то есть 70 тысячных вольта (так называемый потенциал покоя), потому что в клеточной жидкости находится очень много отрицательно заряженных белковых частиц (белковые ионы А", называемые также анионами). При синаптическом возбуждении открываются — начиная с клеточного тела и продолжая до синаптических окончаний — поры в клеточной мембране, которые пропускают положительно заряженные ионы натрия (Na^ назы ваемые катионами) из окружающей клетку межклеточной жидкости. Таким образом, внутри клетки на несколько тысячных долей секунды возникает избыток положительного заряда с напряжением от +30 до +40 милливольт, так называемый потенциал действия.
Потенциал действия снова вызывает у синаптических окончаний выделение нейротрансмиттеров. и возбуждение охватывает следующие клетки (рис. 5 показывает нам в упрощенном виде путь, который проходит нервное возбуждение от рецепторной клетки через нейрон и мотонейрон до органов исполнения — мышц).
Нервные волокна (аксоны) могут иметь разную длину — от нескольких тысячных долей миллиметра до довольно большой длины (некоторые из них простираются от затылочной области до конца спинного мозга, другие — оттуда до пальцев ног). В основном они соединены в толстые пучки, которые обозначаются как периферийные нервы и в своей совокупности образуют периферийную нервную систему. Соединяющиеся друг с другом клеточные тела расположены по группам, в которых находятся вплотную между собой. Эти группы называются ганглиями (нервными сплетениями) — см. рис. 6.

Рис. 6.
|
|
|
В комплексе рептилий и в лимбической системе большинство
— соматических афференций из рецепторов кожи и мышц и
— висцеральных афференций из рецепторов внутренних органов перерабатываются находящимися там регулирующими системами из нервных клеток (способ их действия был подробно обсужден в лекции 2) в управляющие импульсы для всех процессов тела (простые нервные регуляции в форме так называемых рефлекторных дуг мы находим уже в сером веществе спинного мозга; благодаря существующим в нем более коротким проводным путям облегчаются быстрые реакции, например, реактивные).
Через ядра таламуса, которые как главная база регуляции лимбической системы представляют собой своего рода "секретариат" неокортекса, афференций из основных органов чувств, а также отдельные соматические афференций проецируются в соответствующие области коры головного мозга (в таламусе, между прочим, находятся нервные клетки, которые путем спонтанного возбуждения вырабатывают альфа-волны. Так как они играют большую роль в достижении самогипнотического состояния, то таламус в паранормальном отношении занимает ключевую позицию, поэтому о нем мы будем говорить подробнее).
В слуховой и зрительной коре неокортекса перерабатываются слуховые и визуальные, а в чувственной коре — соматические афференций (рис. 10 показывает соотношение величин соматической проекции на кору головного мозга.)
|
|
|
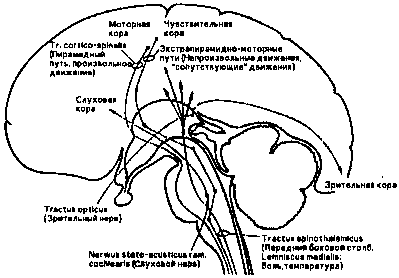
Рис.7. Пять возможных путей нервных волокон.
Небольшая часть обработанной информации передается в сознание для восприятия окружающего мира и тела. Возникающие здесь импульсы действия попадают затем через пирамидный путь (см. рис. 7) — эфферентный волоконный путь — в соответствующие части скелетной мускулатуры, где они вызывают произвольные движения.
Возбуждения, поступающие от рецепторов основных органов чувств, идут через тройничковый нерв непосредственно в мозговой ствол, в то время как рецепторы кожи. мышц и внутренних органов передают свои импульсы возбуждения по афферентным волокнам периферийной нервной системы. (ПНС, рис. 8), нервам серого вещества спинного мозга (рис. 9 и 10). По нервному пучку в заднем отростке спинного мозга — заднему пучку — они достигают также мозгового ствола, откуда попадают через сенсорные клеточные области верхнего мозгового ствола (средний мозг) в лимбическую систему.
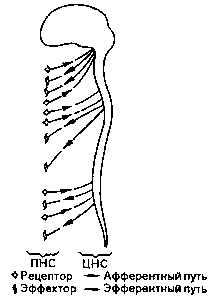 Рис.8. Афферентные и эфферентные нервные пути.
Рис.8. Афферентные и эфферентные нервные пути.
|  Рис.9. Участок спинного мозга с афферентными и эфферентными волоконными путями и с рефлексной дугой. Рис.9. Участок спинного мозга с афферентными и эфферентными волоконными путями и с рефлексной дугой.
|

Рис.10. Ход сенсорных (сенсомоторных) нервных путей от рецепторов тела и головы по позвоночнику. продолговатому телу (Medulla oblongata), стволу мозга и головному мозгу (неокортексу).
Вызов движений через пирамидный путь является, между прочим, единственной возможностью для непарапсихологов вызывать усилием воли процессы в теле, но связанные с этим сопутствующие движения неосознанно управляются чувствительной корой головного мозга и подчиненными областями мозга.
Большинство афференций, которые достигают коры головного мозга, перерабатываются там в ассоциативных полях с помощью накопленных программ и содержательной памяти в управляющие импульсы и поступают через лобную долю, гипоталамус и моторные области ядер верхней части мозгового ствола в боковой и передний отросток спинного мозга.
Естественно, и в подчиненных коре головного мозга мозговых областях тоже происходят процессы переработки, так как имеются прямые соединения между отдельными клеточными ядрами при помощи афферентных и эфферентных нервов. Знание этого не дает ничего дополнительного к пониманию аутогенных процессов, поэтому мы можем отказаться от их более подробного рассмотрения.
Однако, к одной области лимбической системы мы должны будем позже еще вернуться — к гипоталамусу. Эта весящая только 5 граммов мозговая ткань является наиважнейшим для жизни центром регулирования всех вегетативных функций и координации этой вегетативной регуляции с другими видами деятельности организма. Во взаимодействии с гипофизом он является, кроме того. связывающим звеном между центральной нервной системой и эндокринной системой, о которой мы будем говорить в следующей главе из-за важности гормональных процессов регуляции.
Исходящие от спинного мозга эфферентные волокна, которые называются также моторными нервами, распределяются по периферийной нервной системе вплоть до органов действия — эффекторов. В зависимости от их функций они подразделяются на 3 группы:
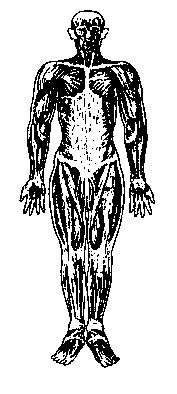
— соматическо-моторные нервы передают приказы центральной нервной системы моторике тела, т.е. тем мышцам, которые отвечают за обеспечение противодействия тела силе тяжести и за его движения (поперечнополосатые или скелетные мышцы — см. рис. 17);
— висцерально-моторные нервы передают управляющие импульсы из вегетативных центров головного мозга гладкой мускулатуре тканей, органов (печени, желудка, кишки, желчного пузыря и почек) и кровеносных сосудов (для их сужения или расширения), а также сердечной мышце:
— секреционно-моторные нервы, наконец, передают поступающие из вегетативных центров мозга приказы о выделении гормонов железам эндокринной системы.
Висцерально- и секреционно-моторные части нервной системы обозначаются как вегетативная нервная система (ВНС. см. лекцию 1). К ней относятся части центральной нервной системы и периферийной нервной системы. Все остальные части нервной системы, преимущественно афферентные и соматическо-моторные, обобщенно называются "соматическая нервная система".
Эффекторами, или органами действия, соматической нервной системы являются скелетные мышцы. Органами действия вегетативной или автономной нервной системы являются все внутренние органы, вся система кровообращения (сердце и сосуды), органы дыхания и все железы.
|
|
|
|
|
|
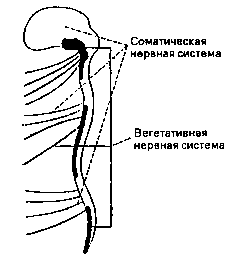 Рис.11.
Рис.11.
| 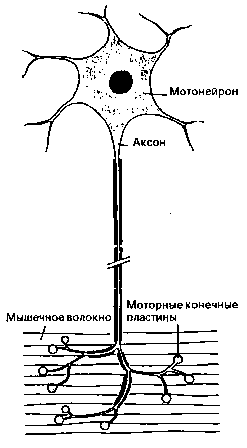 Рис.12. Моторный процесс. Рис.12. Моторный процесс.
|
Итак, вегетативная нервная система управляет прежде всего гладкой мускулатурой внутренних органов, системы кровобращения, органов дыхания и многих желез, в то время как соматическая нервная система передает и перерабатывает информацию от всех органов чувств и управляет скелетной мускулатурой.
Движения гладкой и скелетной мускулатуры достигаются уже упоминавшимся способом:
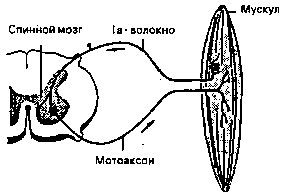
Синаптическое возбуждение переходит от синаптических окончаний нервной клетки на мотонейрон, который вместе с мышечными волокнами, связанными друг с другом моторными конечными пластинами (они соответствуют синаптическим окончаниям нервных клеток), образуют моторное единство (рис. 12).
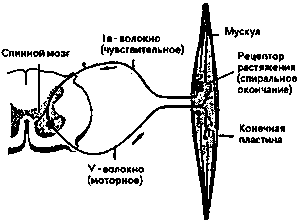
Электрическое возбуждение протекает через аксон мотонейрона и вызывает у моторных конечных пластин выделение молекул трансмиттеров. Последние вызывают сокращение мышечных клеток — и происходит движение. Каждое волокно состоит из многих отдельных мышечных веретен. Рисунок 13 показывает нам одно из них в расслабленном состоянии в сильно упрощенном изображении. Через эфферентный мотоаксон оно связано с нервным узлом (ганглием) серого вещества спинного мозга.
Над моторными конечными пластинами мы видим спиралевидный рецептор растяжения, который, во-первых, через синапсы своего афферентного нервного волокна (называется la волокно) связан с собственным мотонейроном, а, во-вторых, афферентным нервом связан с мозгом. Первая связь, так называемая рефлексная дуга (см. также рис. 9) служит для обеспечения тонуса удержания (состояния напряжения мышцы: при прямом стоянии тонус разгибающих мышц суставов ног и туловища является настолько большим, что точно уравновешивает влияние силы тяжести. При внезапной потере тонуса, например, при нокаутирующем ударе, мы резко падаем на землю).
Рисунок 14 показывает нам сократившееся из-за нервного возбуждения мышечное веретено. Рецептор растяжения передает теперь состояние напряжения мышцы как соматическо-моторную афференцию регулирующим ядрам мозгового ствола и коре головного мозга, где оно перерабатывается дальше.
Этим снова замыкается круг нервной регуляции. Прежде чем перейти к гормональным процессам регуляции. мы в заключение ради наглядности еще раз рассмотрим систему нервов и распределение скелетной мускулатуры, а также схему нервных волокон и автономных центров регуляции.
|
|
|
ГОРМОНАЛЬНЫЕ ПРОЦЕССЫ РЕГУЛЯЦИИ
Наряду с вегетативной нервной системой имеется вторая коммуникативная система для информационного обмена между отдельными органами тела: эндокринная система.
Вегетативная нервная система иннервирует (снабжает поступающими из центральной нервной системы определенными раздражениями конкретные органы) — как мы уже установили — электрическими импульсами в основном гладкую мускулатуру всех органов и систем органов, сердце и железы. Она регулирует важные жизненные функции дыхания, кровобращения, обмена веществ, секреции желез. температуры тела и размножения в тесном взаимодействии с сенсорно-моторной частью нервной системы.
Эндокринная система имеет близкие задачи: вместе с последней она регулирует и координирует функции органов тела. Различие между двумя системами состоит в способе и скорости передачи информации.
В то время как нервная система направляет свои сигналы, закодированные в виде электрических импульсов, по нервным волокнам к отдельным органам, то эндокринная система использует химический "язык молекул" для того. чтобы общаться с исполнительными органами. Нервный метод имеет преимущество в большой скорости. химический же — в том, что хотя он и медленно приводится в действие, однако равномерно действует продолжительное время.
| 
|
| Рис.13. Мышечное веретено в состоянии покоя. | Рис.14. Сократившееся мышечное веретено. |
| 
|
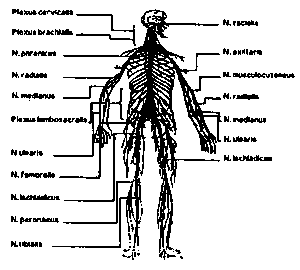
| Рис. 15. Обзор сети нервов. | Рис. 16. Автономные центры регуляции. |
| 
|
| Рис. 17. Скелет с мускулатурой. | Рис. 18. Классификация нервных волокон по происхождению и функциям. |
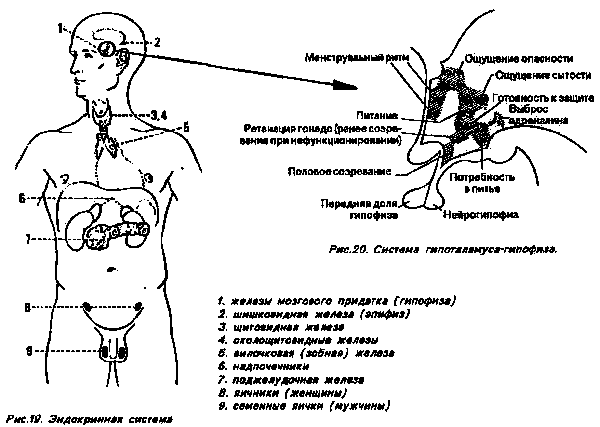
Химические продукты эндокринной системы называются гормонами. Их вырабатывают клетки, которые специализируются исключительно на этом и часто объединены в собственные органы — эндокринные железы (некоторые гормоны вырабатываются также клетками, которые предназначены для других функций — их продукты называют тканевыми гормонами). Рис. 19 показывает нам места расположения эндокринных желез в человеческом теле. Они выделяют в кровь выработанные ими гормоны, которые распределяются ею по всему телу. В местах назначения гормоны вызывают специфические воздействия, то есть такие, которые не могут быть вызваны ни одним другим веществом.
В теле взрослого человека вырабатываются более чем 50 различных гормонов. Они регулируют давление клеток и обмен веществ, обеспеченность энергией и минеральными солями, содержание кальция и сахара в крови, а также функции половых органов (некоторые из продуктов гормонального процесса покидают тело в качестве пахучих веществ с целью вызвать соответствующее возбуждение рецепторных клеток организма другого пола этого же биологического вида).
Импульс для выделения гормонов щитовидная железа, околощитовидные железы, яичники и кора надпочечников (они все называются в совокупности также эффекторными железами) получают от так называемых гландотронных (воздействующих на одну железу) гормонов, которые производятся в железах передней доли гипофиза и выпускаются в кровь (другие производимые в передней доле гормоны — соматотропные гормоны — воздействуют непосредственно на обмен веществ в различных областях тела).
Производство гормонов в передней доле гипофиза стимулируется или тормозится в свою очередь третьей группой гормонов: во многих частях гипоталамуса возникают гормоны "расковывающего действия" и гормоны "сдерживающего действия", которые из гипоталамуса поступают в переднюю долю и там вызывают соответствующие реакции.
Кроме этих гормонов, которые стимулируют или сдерживают производство гормонов в передней доле, нервные клетки гипоталамуса производят также гормоны адиуретин и окситоцин. Они оба передвигаются как нейросекрет вдоль аксонов до задней доли гипофиза (или нейрогипофиза). Оттуда они испускаются в кровь (адиуретин сдерживает выделение воды и значительно повышает кровяное давление; так как он способен вызывать освобождение от страха, то может способствовать повышению концентрации и усилению памяти; окситоцин вызывает испускание молока, если возникает раздражение сосков груди).
Но не только благодаря гормонам "расковывающего или сдерживающего действия", но и за счет нервного возбуждения гипоталамус побуждает другие железы к производству и выделению гормонов. Нервным путем (латинское:
Fasciculus longitudinalis dorsalis) он связан с ядрами симпатической части вегетативной нервной системы в спинном мозге и регулирует через них функции поджелудочной железы и внутренних тканей надпочечников.
Итак, в целом система гипофиз-гипоталамус представляет из себя со всем многообразием задач по управлению и регуляции, которые нам еще раз ясно показывает рисунок 22, важнейший направляющий центр всей нервной системы.
Гипоталамус связан через таламус с лимбической системой. через лобный кортекс — с большим мозгом. Обе мозговые области передают ему постоянно афферентные сведения о состоянии окружающего мира и тела (фактическое значение), которые он сравнивает с накопленными в
нем заданными значениями. Если он устанавливает какое-либо отклонение, то приводит в действие нервные и гормональные ответные реакции (некоторые из регулирующих центров внутри гипоталамуса мы видим на рис. 20).
Так гипоталамус во взаимодействии с остальной нервной системой обеспечивает последовательное приспособление тела к изменяющимся условиям внешнего мира. Ярким примером способа действия этих механизмов приспособления являются соматические реакции на стресс. Под стрессом понимаются тяжелые внешние нагрузки на организм: повреждения, крайние жара или холод, тяжелая физическая работа или ограниченность в движении, а также резкое превышение или снижение порога раздражения органов чувств (звук), внутренние нарушения (инфекционные болезни, очень сильные эмоции: гнев. радость, забота и ненависть).
На первой фазе подобного интенсивного нарушения внутреннего равновесия, называемого шоком, гипоталамус через лимбическую систему возбуждает центральную нервную систему, которая через свою вегетативную часть приводит в действие первые нервные реакции, такие как
повышение частоты ударов сердца и тонуса мышц (для углубления дыхания и сужения периферийных кровеносных сосудов).
Параллельно с этой серией импульсов вегетативной нервной системы гипоталамус вызывает и возбуждение эндокринной системы: через Fasciculus longitudinalis dorsalis (ФЛД) он направляет внутренним тканям надпочечников (ВТН) командный импульс на выделение гормонов адреналина и норадреналина. Адреналин усиливает превращение энергии, мобилизует запасы сахара и вызывает повышенную частоту ударов и силу ударов сердца. Норадреналин вызывает сужение поверхностных сосудов (таким образом кровь вытеснятся из периферии тела, что особенно важно при повреждениях, а жизненно важные органы — сердце, мозг и печень могут быть за счет этого лучше обеспечены).
В то время как шоковая фаза должна обеспечить сохранение жизни при обширных повреждениях, организм готовится в наступающей в след за ней "антишоковой" фазе к самозащите и восстановлению. Для этого вегетативная регуляция приводит в действие процессы, обеспечивающие поступление энергии. В частности, гипоталамус побуждает (наряду с выделением адреналина и норадреналина) к производству "расковывающего" гормона СРН (см. рис. 21). Последний вызывает в гипофизе выделение гормона АСТН, который попадает в кровь и побуждает кору надпочечников произвести и выделить гормоны кортизол и Тз.

Тз поддерживает ускоренный обмен веществ, под воздействием адреналина кортизол производит из аминокислот сахар, поддерживает воздействие адреналина, направленное на усиление сердечной деятельности, и общее сосудосужающее воздействие норадреналина, усиливает выделение желудочного сока и оказывает антивоспалительное воздействие.
Нарушения вегетативного и гормонального внутриорганического равновесия при повторяющемся или постоянном стрессе могут вызвать разнообразные органические болезни или способствовать инфекционным заболеваниям. Сутью этих так называемых психосоматических болезней и возможностями их лечения путем самовнушения (духовного самоизлечения) мы однако займемся только в следующей лекции. А пока еще одно замечание.
Только благодаря телесному и духовному расслаблению, которое оказывает успокаивающее воздействие на всю нервную систему и, значит, на гипоталамус, постепенно проходят сами по себе или совершенно исчезают многие психосоматические нарушения и болезни. Лучшим доказательством этого являются многие сообщения, присланные в ИППН уже по истечении всего нескольких месяцев изучения парапсихологии.
Например, госпожа Гудрун А. из Италии пишет:
"... Исключительно содержательные лекции помогли мне изменить мои взгляды на жизнь и снова восстановить мое душевное равновесие, которое я потеряла из-за чрезвычайно тяжелой болезни. И не только это: я смогла избавиться от своего недуга, так что сейчас я чувствую себя. как только что родившаяся."

Рис. 22. Схема воздействия системы гипоталамус - гипофиз.
Естественно, здесь речь идет не о сознательной форме самоизлечения, а только о положительном — хотя и неоценимом — побочном эффекте расслабления, которое способствует в первую очередь достижению состояния самогипноза. А теперь мы хотим подробнее рассмотреть расслабление и его психологически-физиологические основы.
ТЕЛЕСНОЕ И ДУХОВНОЕ РАССЛАБЛЕНИЕ
По всей своей длине мозговой ствол пронизан вертикально расположенной сетью нервов, которая связывает чувствительные (афферентные) с соматическо- и висцерально-моторными мозговыми нервными ядрами — это ретикулярная (сетчатая) система. Многозвенными цепями нейронов она к тому же вверху связана с лимбической системой, сзади — со средним мозгом, а внизу — с моторными клетками переднего отростка спинного мозга.
Рисунок 23а показывает нам положение ретикулярной системы при поперечном разрезе мозга, а схема на рис. 236 — поперечные связи с другими областями мозга (в сильно упрощенной форме).
Ретикулярная система регулирует бодрость всей нервной системы и таким образом ответственна за силу всех нервных состояний возбуждения. Исходя из ее функций. ее проще всего можно сравнить с радиоприемником, в котором поступающие с антенны слабые электрические импульсы в зависимости от установленной в приемнике громкости в большей или меньшей степени становятся слышны как акустические сигналы.
"Антенными импульсами" ретикулярной нервной системы являются возбуждения рецепторов, которые передаются ей через ответвления афферентных волоконных путей в среднем мозгу, а также возбуждения из лимбической системы, гипоталамуса и лобного кортекса.
В ретикулярной системе поступающие импульсы возбуждения усиливаются в большей или меньшей степени и передаются в форме неспецифических (не четко очерченных) активирующих импульсов моторным областям ядер верхнего мозгового ствола (для активизации вегетативных функций), переднему отростку спинного мозга (для активации совершающейся там координации рефлексов с процессами движения), а также коре головного мозга (для осознанного восприятия и переработки раздражителей).
Как происходит активация коры головного мозга, мы хотим продемонстрировать на одном примере: по специфическим нервным путям внешний раздражитель чувств — "звонок будильника" — попадает от ушей в особую часть коры головного мозга, которая регистрирует и перерабатывает эту информацию, так сказать, в форму "будильник звонит". Одновременно этот акустический раздражитель по ответвлениям стимулирует ретикулярную систему, которая затем сама по себе активирует по неспецифическим путям весь неокортекс с тем конечным результатом, что мы просыпаемся и начинаем осознанно думать: "Я должен вставать".
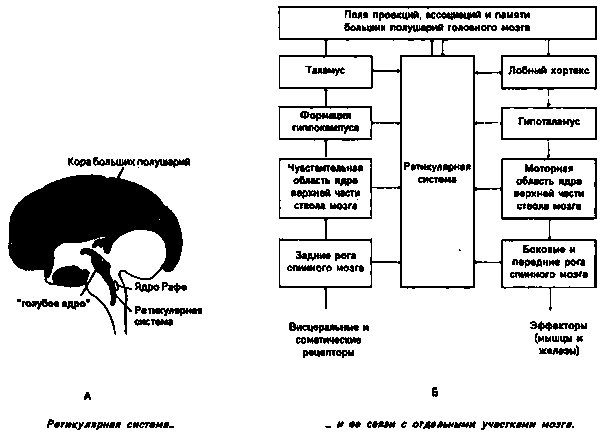
Рис.23.
Итак, каждому осознанному действию неокортекса и каждому возрастанию моторных и вегетативных функций необходима определенная мера активизации через ретикулярную систему!
Используя электроэнцефалографию (ЭЭГ, см. лекцию 1). можно сделать активность коры головного мозга видимой (на электроэнцефалограмме). Для этого на черепе и на мочках ушей прикрепляются электроды, направляющие часть нервных токов в усилитель. Последний в свою очередь связан с самописцем, который изображает последовательность импульсов и потенциал (электрическое напряжение) в виде зубчатой кривой на рулонной бумаге.
Если мы находимся в нормальном (и даже возбужденном, нервном или испуганном) состоянии бодрствования с напряженным вниманием, неподвижны, то наша ЭЭГ покажет преимущественно бета-волны: Их частота большей частью не превышает 20 — 25 колебаний в секунду, а их амплитуда — от 2 до 20 микровольт.
Рис.24. Схема подключения электродов при ЭЭГ.
При визуальной или слуховой стимуляции коренным образом изменяется волновая картина этой так называемой "фоновой активности" мозга. Некоторые верхние точки достигают амплитуды до 200 микровольт.
Если же импульсы активации из ретикулярной системы ослабевают, то заметно снижается "фоновая активность" и уступает место типу волн, который в последнее время стал модным явлением: альфа-волнам с частотой от 8 до 12 герц и амплитудой от 5 до 100 микровольт.
Почти все авторы книг и учебных пособий о способах расслабления, аутогенном тренинге, трансцендентальной медитации, йоге или альфа-тренинге называют "производство" альфа-волн в мозгу предпосылкой для достижения того самогипнотического состояния, которое позволяет успешную работу по самовнушению. При этом они игнорируют — сознательно или по незнанию — важные нейрологические условия.
Альфа-волны не "производятся" в головном мозгу. Они скорее всего возникают в так называемых "клетках-лидерах" таламуса и синхронизируют нейронную деятельность головного мозга, как только последний путем ликвидации раздражителей закрытием глаз (доля визуальных раздражителей во всем объеме раздражении, которые достигают коры головного мозга, составляет около 90%!), успокаивает лимбическую систему и ослабленная активация ретикулярной системы не может больше вызывать возбуждений.
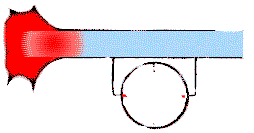
Рис.25. Возникновение "температурных кривых" ЭЭГ на примере отвода потенциала действия с мембраны одного нейрона.Между двумя электродами проявляется разница напряжений (движение кривой вверх или вниз) тогда. когда двигающаяся слева направо (положительная) волна возбуждения задевает только один из них (голубые участки клетки свидетельствуют о потенциале покоя, а красные — о потенциале действия).
Достигнутое таким образом расслабление, уютно-мечтательное, но и творческое состояние бодрствования не имеет ничего общего с самогипнотическим состоянием:
лобный кортекс проявляет все еще свою типичную бета-активность, и если открыть глаза, то альфа-волны исчезают снова в полях проекций, воспоминаний и полях ассоциаций неокортекса, уступая место бета-волнам.
Даже в состоянии пустоты мыслей, когда альфа-волны могут распространяться в лобном кортексе — местонахождении сознания, еще не возможна эффективная связь с бессознательными областями мозга. Ведь они "говорят" преимущественно на другом "языке" — на серийных импульсах от 4 до 7 в секунду и амплитуде от 5 до 150 микровольт, т.е. на так называемых тета-волнах.
Только тогда, когда становится возможным сознательно производить тета-волны в лобном кортексе, достигается состояние самогипнотического сна и появляется доступ к бессознательным областям центральной нервной системы.
Наилучшее и наиболее зримое доказательство правильности этого вывода мы получаем тогда, когда просыпаемся посреди ночи после яркого, живого, подчеркнуто чувственного и причудливого сновидения, чье содержание не должно совпадать с тем. что возможно в "реальном мире".
Во время этих сновидений, появляющихся в так называемом быстром сне (из-за быстрых движений глаз — признака стадии сновидений), которые четко отличаются от второго типа сновидений в процессе медленного сна со скорее рациональным, реалистичным и схожим с мыслями содержанием, происходит коммуникационный обмен между сознанием и подсознанием: архетипы, т.е. коллективные
содержания подсознания, перерабатываются вместе с личностным содержанием памяти в сновидения — и все это время ЭЭГ фиксирует тета-волны!
Чтобы понимать связанные с достижением самогипнотического сна (который идентичен быстрому сну в том, что вызывает тот же волновой образец на ЭЭГ и допускает связь между сознанием и подсознанием — но при том, что здесь она протекает не "случайно", а направляется сознательно) нейронные процессы в центральной нервной системе, мы хотим разобраться с тем, что происходит, когда мы спим.
Вхождение в состояние сна производится нервным центром в мозговом стволе — ядром Рафе (см. рис. 23а), который — возвращаясь к нашему первоначальному сравнению с радиоприемником — играет роль "регулятора силы звука" для ретикулярной системы. Через соответствующие командные импульсы ядро Рафе уменьшает ее активность настолько, что она больше не передает никаких активирующих импульсов центральной нервной системе и приглушает афферентные рецепторные возбуждения до минимальных величин, необходимых для спокойного протекания жизненных процессов.
Поэтому при засыпании снижается не только степень бодрствования коры головного мозга, но и расслабляется тонус мышц, успокаивается дыхание, замедляются сердечные удары.
До приблизительно одного часа после засыпания постоянно снижается частота мозговых волн: преимущественные частоты, которые проявляются при этом на ЭЭГ. ступенчато меняют свою градацию от альфа-, через тета-, до дельта-волн. Стадия медленного сна достигнута, если проявляются только медленные (0,3—4 колебаний в секунду) и большие (амплитуда в 20—200 микровольт) дельта- волны.
Спустя приблизительно 1,5 часа после засыпания в действие вступает второй нервный центр: голубое ядро. которое находится у центров рефлексо-подобных регуляций в среднем мозгу (см. рис. 23а), оно возбуждает "клетки-лидеры" для тета-волн в перегородке — тонкой связующей преграде между двумя половинами переднего мозга (т.е. между правым и левым лобным кортексом. таламусом и гипоталамусом). Исходя из этой перегородки, тета-волны синхронизируют возбуждение остальных нервных клеток в мозгу и возникает таким образом единое возбуждение всего мозга (сдерживающее воздействие ретикулярной системы этим частично устраняется: вместо нее текущее поведение сдерживают теперь импульсы, которые из голубого ядра попадают через нервный пучок в лимбическую систему).
Сознание и подсознание "говорят" теперь на одном языке, между ними возможен беспрепятственный обмен информацией.
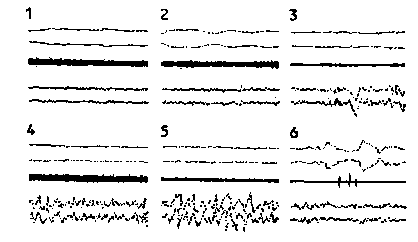
Рис.26. Протекание сна от состояния бодрствования (1) через фазы медленного сна (2—5) до быстрого сна (б). Обе верхние кривые показывают движения глаз. средняя — тонус подбородочной мышцы, нижние ЭЭГ.
|
|
|
