 |


22.Врожденный иммунитет и система распознавания
|
|
|
|
22. Врожденный иммунитет и система распознавания
Врождённый иммунитет является присущей каждому организму с рождения, генетически закреплённой способностью противостоять инфекции. Это передовая линия обороны организма против патогенов, пытающихся проникнуть или уже проникших в покровные ткани или внутреннюю среду. Врождённый иммунитет срабатывает мгновенно или в течение первых нескольких часов после контакта с патогеном (возбудители, выделяемые ими токсины и другие чужеродные молекулы). Он включает четыре основных уровня защиты: анатомический, физиологический, фагоцитарный и воспалительный - покровные ткани, фагоциты, микробоцидные гуморальные вещества (протеазы, сильные окислители и свободные радикалы, продуцируемые фагоцитами, эндогенные противомикробные пептиды и др. ), сосудистые реакции. Первичные рецепторы врождённого иммунитета - это молекулы многоклеточных, позволяющие «считывать» эволюционную память - информацию о том, чем отличаются микроорганизмы от собственных клеток.
Эти рецепторы способны распознавать консервативные молекулярные структуры - РАМР (Pathogen-Associated Molecular Patterns), характерные для групп сходных микроорганизмов. В настоящее время постоянно открывают новые патогенраспознающие рецепторы врождённого иммунитета. К ним относят мембраносвязанные паттернраспознающие рецепторы (Pattern Recognition Receptors, PRR), а также растворимые рецепторы - ряд белков сыворотки крови: C-реактивный белок (СРБ), маннозосвязывающий лектин (MBL - Mannose-Binding Lectin), компоненты комплемента.
23. Адаптивный иммунитет и система распознавания
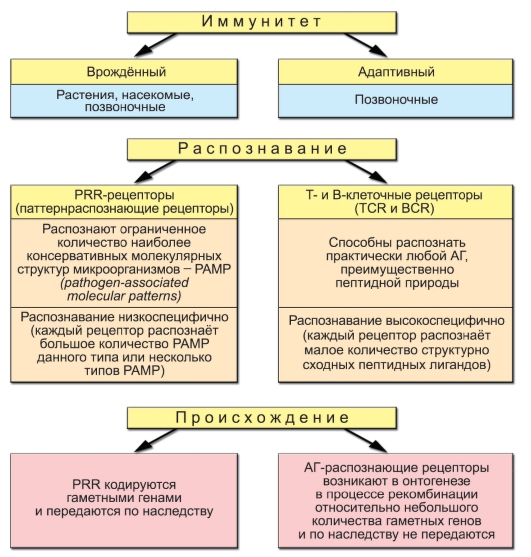 Собственные возможности клеток врождённого иммунитета санировать организм от проникшего патогена часто недостаточны. Множество патогенов приспособилось выживать в присутствии факторов врождённой резистентности к инфекциям. Именно поэтому в процессе эволюции, начиная с челюстных рыб, к врождённому иммунитету добавился адаптивный иммунитет - специфический. Материальные носители адаптивного иммунитета - лимфоциты. Уникальное и отличительное свойство лимфоцитов как множества клеток - способность распознавать почти неограниченное (1018) разнообразие молекулярных объектов - антигенов. Лимфоциты характеризуются экспрессией Т-клеточных (TCR) или В-клеточных (BCR) рецепторов, распознающих только одну антигенную детерминанту либо небольшое число структурно очень близких детерминант, и поэтому, в отличие от PRR, обладающих высокой специфичностью (рис. 1-1).
Собственные возможности клеток врождённого иммунитета санировать организм от проникшего патогена часто недостаточны. Множество патогенов приспособилось выживать в присутствии факторов врождённой резистентности к инфекциям. Именно поэтому в процессе эволюции, начиная с челюстных рыб, к врождённому иммунитету добавился адаптивный иммунитет - специфический. Материальные носители адаптивного иммунитета - лимфоциты. Уникальное и отличительное свойство лимфоцитов как множества клеток - способность распознавать почти неограниченное (1018) разнообразие молекулярных объектов - антигенов. Лимфоциты характеризуются экспрессией Т-клеточных (TCR) или В-клеточных (BCR) рецепторов, распознающих только одну антигенную детерминанту либо небольшое число структурно очень близких детерминант, и поэтому, в отличие от PRR, обладающих высокой специфичностью (рис. 1-1).
|
|
|
Адаптивный (приобретённый) иммунитет формируется в течение жизни индивидуума. Активно приобретённый иммунитет - состояние невосприимчивости к инфекции после перенесённого инфекционного заболевания или после вакцинации (сам организм вырабатывает соответствующие антитела). Пассивно приобретённый иммунитет - состояние невосприимчивости к инфекции в результате поступления в организм уже готовых антител от матери или в результате инъекции (сам организм эти антитела не вырабатывает).
24. Принципы передачи сигналов
В большинстве случаев передача сигнала внутри клетки представляет собой цепь последовательных биохимических реакций, осуществляемых ферментами, часть из которых активируется вторичными посредниками. Такие процессы обычно являются быстрыми: их продолжительность — порядка миллисекунд в случае ионных каналов и минут — в случае активации протеинкиназ и липид-опосредованных киназ. Однако в некоторых случаях от получения клеткой сигнала до ответа на него могут проходить часы и даже сутки (в случае экспрессии генов). Пути передачи сигнала, или сигнальные пути, часто бывают организованы как сигнальные каскады (англ. signal cascade): количество молекул белков и других веществ, принимающих участие в передаче сигнала, возрастает на каждом последующем этапе по мере удаления от первоначального стимула. Таким образом, даже относительно слабый стимул может вызывать значительный ответ. Это явление называется амплификацией сигнала.
|
|
|
В зависимости от функций первичные посредники могут быть разделены на несколько групп:
· гормоны
· цитокины
· нейротрансмиттеры
· факторы роста
Рецепторы[править | править код]
Получение клеткой сигнала от первичных посредников обеспечивается особыми белками-рецепторами, для которых первичные посредники являются лигандами. Для обеспечения рецепторной функции молекулы белков должны отвечать ряду требований:
Клеточные рецепторы делятся на следующие классы:
· мембранные
o рецепторные тирозинкиназы
o рецепторы, сопряжённые с G-белками
o ионные каналы
· цитоплазматические
· ядерные
Мембранные рецепторы распознают крупные (например, инсулин) или гидрофильные (например, адреналин) сигнальные молекулы, которые не могут самостоятельно проникать в клетку. Небольшие гидрофобные сигнальные молекулы (например, трийодтиронин, стероидные гормоны, CO, NO) способны проникать в клетку за счёт диффузии. Рецепторы таких гормонов обычно являются растворимыми цитоплазматическими или ядерными белками. После связывания лиганда с рецептором информация об этом событии передаётся дальше по цепи и приводит к формированию первичного и вторичного клеточного ответа[2].
Механизмы активации рецепторов [править | править код]
Если внешняя сигнальная молекула воздействует на рецепторы клеточной мембраны и активирует их, то последние передают полученную информацию на систему белковых компонентов мембраны, называемую каскадом передачи сигнала. Мембранные белки каскада передачи сигнала подразделяют на:
· белки-преобразователи, связанные с рецепторами
· ферменты-усилители, связанные с белками-преобразователями (активируют вторичные внутриклеточные посредники, переносящие информацию внутрь клетки).
Так действуют рецепторы, сопряженные с G-белками. Другие рецепторы (ионные каналы, рецепторы с протеинкиназной активностью) сами служат умножителями.
|
|
|
Вторичные посредники[править | править код]
Вторичные посредники (англ. second messenger) — это низкомолекулярные вещества, которые образуются или высвобождаются в результате ферментативной активности одного из компонентов цепи передачи сигнала и способствуют его дальнейшей передаче и амплификации[2]. Вторичные посредники характеризуются следующими свойствами: имеют небольшую молекулярную массу и с высокой скоростью диффундируют в цитоплазме; быстро расщепляются и быстро удаляются из цитоплазмы. Ко вторичным посредникам относятся:
· ионы кальция (Ca2+);
· циклический аденозинмонофосфат (цАМФ) и циклический гуанозинмонофосфат (цГМФ)
· инозитолтрифосфат
· липофильные молекулы (например, диацилглицерол);
· оксид азота (NO) (эта молекула выступает и в роли первичного посредника, проникающего в клетку извне).
Иногда в клетке образуются и третичные посредники. Так, обычно ионы Ca2+ выступают в роли вторичного посредника, но при передаче сигнала с помощью инозитолтрифосфата (вторичный посредник) выделяющиеся при его участии из ЭПР ионы Ca2+ служат третичным посредником.
Передача сигнала предполагает примерно следующую схему:
1. взаимодействие внешнего агента (стимула) с клеточным рецептором,
2. активация эффекторной молекулы, находящейся в мембране и отвечающей за генерацию вторичных посредников,
3. образование вторичных посредников,
4. активация посредниками белков-мишеней, вызывающих генерацию следующих посредников,
5. исчезновение посредника.
|
|
|
