 |


PI3-киназная сигнальная система, механизмы включения и выключения этого сигнального каскада.
|
|
|
|
Свое название PI3-киназа получила согласно своей субстратной специфичности. Этот фермент представляет пример липидно-белковой киназы, способной фосфорилировать и низкомолекулярные липиды, и серин/треониновые аминокислотные остатки в составе белков. Как основной субстрат PI3-киназа использует фосфатидилинозитол (PI) и переносит остаток фосфата с молекулы АТФ в третье положение инозитольного кольца этого фосфолипида. Различные изоформы PI3-киназ фосфорилируют как сам фосфатидилинозитол (PI), так и его фосфопроизводные – монофосфо-PI или бисфосфо-PI. Некоторые исследователи рассматривают эти липидные молекулы как вторичные посредники, которые локализованы в липидном бислое и усиливают сигнал благодаря тому, что образуются в большом количестве в ходе ферментативной реакции.
PI3-киназный сигнальный каскад активируется большинством тирозинкиназных и многими семидоменными рецепторами. Всего известно несколько изоформ PI3-киназ, которые разделяются на три класса, обозначаемых как I, II и III. PI3-киназы класса I являются основными участниками сигнальных каскадов, активируемых поверхностными рецепторами. Они образуют фосфатидилинозитол-3,4,5-трисфосфат (PIP3) из фосфатидилинозитол-4,5-бисфосфата (PI-4,5-P2). Ферменты двух других классов фосфорилируют, в основном, фосфатидилинозитол до PI(3)P и функционируют во внутренних компартментах клетки, регулируя трафик эндосом и лизосом. PI3-киназы класса I являются гетеродимерами, в состав которых входит одна из 4-х возможных (α, β, γ и δ) каталитических субъединиц весом 110 кДа и одна из большого набора регуляторных субъединиц.
Общая схема PI3-киназного каскада представлена на рис. 83. С рецепторами связываются регуляторные субъединицы PI3-киназы р85, которые далее связывают и активируют каталитические субъединицы PI3-киназы. При этом Рецепторы факторов роста, в том числе Рецептор PDGF в фибробластах и Рецептор EGF в эпителиальных клетках, дополнительно используют малую ГТФ-азу Ras для полноценной активации PI3-киназного комплекса. Весь процесс активации происходит около мембраны в связанном с рецептором состоянии. Для повышения эффективности используется принудительная локализация этого комплекса на мембране за счет множественных взаимодействий с рецептором и липидных якорей в молекуле Ras. Рисунок 83:
|
|
|
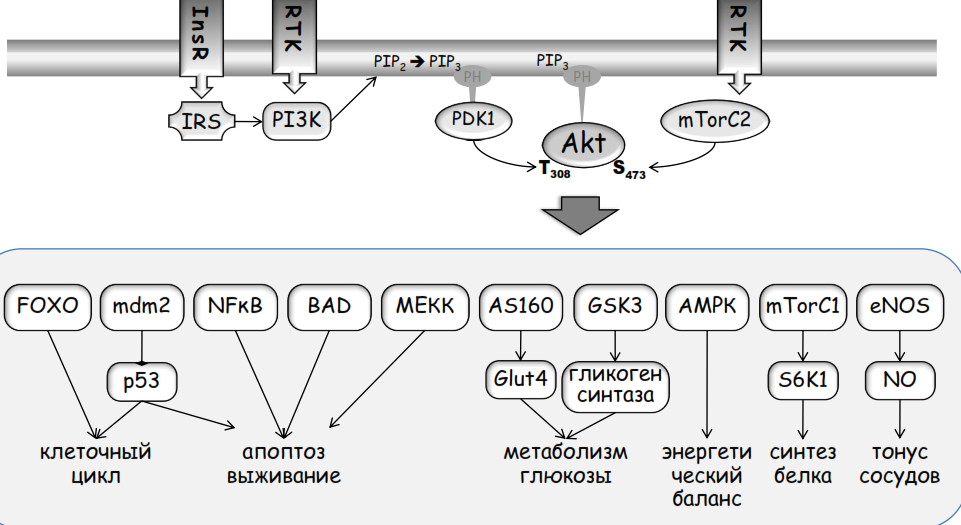
PI3-киназа служит одним из главных эффекторов инсулинового рецептора (InsR на рис. 83). Однако в этом случае за прямую активацию отвечает субстрат инсулинового рецептора – белок IRS (англ. insulin receptor substrate) (рис. 83). Наличие IRS является отличительной чертой инсулиновой сигнализации. Именно IRS опосредует передачу сигнала от рецептора ко всем нижестоящим мишеням, сохраняя общий принцип передачи сигнала с помощью тирозинового фосфорилирования. Фосфорилированный рецептором по остаткам тирозина IRS служит посадочной площадкой для сборки всех последующих инсулин-зависимых сигнальных каскадов, включая PI3-киназу. IRS привязывает этот процесс к мембране, поскольку содержит РН-домен, взаимодействующий с фосфоинозитидами.
На тирозинкиназных рецепторах (RTK) собираются PI-киназные комплексы, несущие α, β или δ каталитические субъединицы PI3-киназы. Рецепторы, сопряженные с G-белками (GPCR), активируют PI3Kγ изоформу PI3-киназы. В этом случае за активацию отвечает комплекс βγ-субъединиц гетеротримерных G-белков, но детали процесса пока малопонятны.
Заякоренная рецептором на мембране PI3-киназа фосфорилирует PIP3 до PIP2. Обратную реакцию гидролиза PIP3 катализируют фосфатазы PTEN (англ. phosphatase and tensin homolog) и SHIP (англ. Src-homology 2 (SH2)-containing phosphatase – фосфатаза, содержащая SH2-домен -домен Src киназной гомологии 2 класса (порядка); домен адаптерных взаимодействий, гомологичный одному из доменов Src кинзы, связывает фосфорилированные тирозины (Src Homology 2)). PTEN дефосфорилирует PIP3 по 3 положению гидроксильной группы (превращая его в PI-4,5-P2), а SHIP – по 5 положению (при этом образуется PI-3,4-P2). SHIP не полностью терминирует сигнал от рецептора, так как некоторые белки способны связываться и активироваться не только PIP3, но и PI-3,4-P2. Например, протеинкиназа PKB/Akt взаимодействует с обоими липидами. Продукт активности SHIP способен вызывать клеточные ответы как сходные с PIP3, так и отличные от него. Фосфатаза PTEN является опухолевым супрессором и ее нарушения часто встречаются при опухолевых трансформациях, так как ведут к повышенной и неуправляемой миграторной и пролиферативной активности клеток. Функции PTEN и SHIP в регуляции направленной миграции клеток связаны с поляризацией цитоскелета и обособлением лидирующей мембраны, преимущественно содержащей актин, и задней части клетки, обогащенной миозином.
|
|
|
Образующийся в мембране PIP3 связывает белки, имеющие в своем составе РН-домен (рис. 83). Многие белки, содержащие этот домен, активируются именно в результате связывания с PIP3. К ним относятся факторы обмена ГДФ на ГТФ для малой ГТФазы Rac, и некоторые цитоплазматические тирозинкиназы. Активация Rac имеет прямое отношение к миграции клеток, так как это основной регулятор актиновой динамики и образования псевдоподий. Таким образом, Rac и активирующий его PI3-киназный сигнальный каскад являются основными регуляторами поляризации клеток.
Другие белки не активируются PIP3, но просто связываются с мембраной, обогащенной этим липидом. Собираясь в одном месте мембраны, эти белки получают возможность взаимодействовать и активировать друг друга, тогда как, находясь в цитоплазме, они пространственно разнесены. Например, посредством своих РН-доменов с мембранным PIP3 связываются протеинкиназа В (Akt) и активирующая ее фосфоинозитид-зависимая киназа (англ. phosphoinositide-dependent kinase-1, PDK1). Киназа PDK1 конститутивно активна, но, только связавшись с мембраной, оказывается вблизи своего субстрата PKB/Akt. Мишени PKB/Akt в клетке многочисленны (рис. 83), и их список продолжает увеличиваться. Они контролируют жизненно важные функции клетки. Это не только движение и поддержание метаболического баланса, но и стратегический выбор дальнейшей судьбы клетки. Именно через РКВ/Akt PI3-киназный каскад реализует действие гормонов, факторов роста и цитокинов в отношении роста, деления или дифференцировки, поддержания жизнеспособности или клеточной смерти, активного движения или метаболического баланса клеток. Дополнительные рисунки:
|
|
|




|
|
|
