 |


DXM против болезней и различных состояний 10 глава
|
|
|
|
9.3. Почему у DXM существуют плато?
9.3.1. Плато 1-3: Совокупность рецепторов
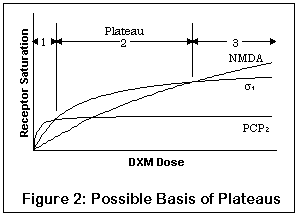
На рисунке изображен вероятный эффект совокупности рецепторов. Заметьте - это качественный рисунок, а не количественный; настоящие уровни насыщения различных рецепторов для данной дозы DXM все еще практически неизвестны. Кривые означают уровни насыщения в PCP2, сигма1 и сайте связывания в открытом канале NMDA, полное насыщение соответствующего рецептора обозначено "уплощением" кривой. Ось Х -концентрация DXM; Y - процент насыщения рецепторов.
Между прочим, если вы знакомы с вещами подобного рода, вам может быть удобнее пользоваться координатами Lineweaver-Burk, но для непосвященных это может оказаться смущающим.
Из-за повышенного сродства к PCP2, сигма1 и NMDA-рецепторам соответственно (сигма2 не представлен), малая доза будет оказывать пропорционально больший эффект на PCP2-рецепторы, в то время как увеличение дозы приведет к насыщению этих рецепторов. Прием большего количества DXM не изменит уровень насыщения PCP2 существенным образом, а окажет влияние на другие рецепторы.
Более того, более слабые эффекты воздействия на PCP2-рецепторы может быть почти полностью сглажены эффектами сигма1 и NMDA-рецепторов (различные уровни насыщения показывают это). Это совершенно разумно, поскольку и сигма1 и NMDA-активность выявляют довольно сильное изменение поведения, последний в большей степени, чем первый.
Итак, первое плато может соответствовать доминированию PCP2-активности с некоторой сигма-активностью и небольшим эффектом NMDA-блокады; второе плато - сигма и некоторым NMDA-эффектам; третье - глубокой NMDA-блокаде.
Это, несомненно, упрощение, и не принимает во внимание индивидуальные отклонения в рецепторах. Некоторые люди могут иметь генетические варианты рецепторов, к которым DXM имеет более сильное (или слабое) сродство. Человек с PCP2-рецепторами, сильно связывающимися с DXM, может наслаждаться стимулирующими эффектами гораздо больше других.
|
|
|
К тому же, ионные каналы и сигма2-рецепторы в рисунке не указаны. Однако они, несомненно, вносят свой вклад, и некоторые люди утверждают, что есть плато и между первыми тремя. Они могут быть неярко выраженными и незаметными для большинства.
9.3.2. Четвертое Плато: Отключение чувств

А что происходит на четвертом плато? Ну, вот еще один маленький рисуночек, который, надеюсь, прояснит все. Это рисунок нейросети; точки - нейроны, линии - их соединения. Как и все в мозгу, эта нейросеть сильно связана. Проценты - количество оставшихся функциональных связей.
Когда достаточное число NMDA-рецепторов блокировано, нейрон может потерять большую часть входящей информации от другого нейрона и соединение эффективно нарушается. Вначале это не причиняет неудобств, поскольку нейроны и связи нейронов еще достаточно плотны и другие нейроны могут взять на себя дополнительную нагрузку (хотя результатом может стать более медленная и менее аккуратная работа нейросети). В некоторый момент, достаточная связанность теряется и нейросеть уже больше не может функционировать.
Сравните это с диссоциацией четвертого плато. На некотором уровне, некие части мозга (вероятно, гиппокамп) теряет способность к функционированию настолько, что больше не может работать как единый блок. Сенсорный процессинг останавливается, и необработанная входящая сенсорная информация не может быть преобразована в соответствующе проанализированный (parsed) выходной поток. Сознание остается без реальной информации; вместо этого образуются хаотические нестабильные мыслеформы (паттерны). Следовательно - диссоциативная анестезия.
|
|
|
С другой стороны, может случиться так, что порог четвертого плато достигается, когда сенсорная информация тормозится достаточно для того, чтобы нейросигналы в лимбических областях отражались назад (feed back) на вход без существенного подавления со стороны входящей сенсорной информации. Эта петля обратной связи продолжает возмущать нейропаттерны до тех пор, пока они не становятся оторванными от состояния человека или его воспоминаний.
9.4. Почему это так сложно?
DXM сам по себе очень сложное вещество; большинство наркотиков связываются с одним или двумя рецепторами (по крайней мере, с одним классом рецепторов). Рекреационное злоупотребление им, известное уже долгие годы, еще не изучалось должным образом, он может воздействовать по-разному на разных людей. Рецепторы и сайты связывания, на которые он воздействует - сигма, NMDA и PCP2 - недавние открытия. Все это приводит к сложному и плохо понимаемому наркотику.
Кроме того, мозг - это сложная система, и, по большому счету, мы ничего не понимаем в его работе. Основы нейропередачи (нейротрансмиссии) вроде бы поняты, но остается много вопросов. Никто не знает, почему так много нейромедиатров или подтипов рецепторов. Еще хорошо не изучены системы вторичных посредников большинства рецепторов. Большинство событий внутри нейронов происходит путем изменений в экспрессии генов, а это отдельная тема, о которой мало известно.
Повторю часто цитируемые (и правильные) слова: если бы наш мозг можно было бы просто понять, мы бы были такими примитивными, что не смогли бы его понять. Я верю, что, в конце концов, мы узнаем, как работает мозг, но это случится не при моей жизни. До тех пор мы просто пытаемся догадаться, шаря в темноте.
9.5. Фармакокинетика: Как метаболизируется DXM
DXM, как соль гидробромида, быстро абсорбируется из желудочно-кишечного тракта; в течение 30 минут он может войти в кроветок. Соединение полистирекса предназначено для продолжительной абсорбции, и его полный переход в кровь может занять 6-8 часов.

DXM метаболизируется двумя путями, оба из них приводят к одному - 3-гидроксиморфинану (3ГМ, 3HM). Первый путь исходит из DXM к DXO (декстрорфан), а затем к 3HM; второй из DXM к 3-метоксиморфинану (3ММ), а затем к 3HM. Большая часть DXM (до 90%) метаболизуется через DXO у нормальных индивидуумов.
|
|
|
DXM преобразуется в DXO печеночным ферментом, называемым цитохром P450-2D6 (дебрисохин 4-гидроксилаза, debrisoquine 4-hydroxylase). Примерно у 7% белых и 0,5% азиатов версия этого фермента работает крайне (до 70 раз медленнее) неэффективно и не может метаболизировать DXM в DXO должным образом. От 0.5 до 2% популяции имеют многочисленные копии гена P450, что приводит к очень быстрому метаболизму DXM в DXO (155). После превращения в DXO, ферменты P450-3A4 и P450-3A5 перобразовывают DXO в 3-гидроксиморфинан.
Другой путь ведет сначала к 3-метоксиморфинану (посредством ферментов P450-3A4 и P450-3A5), а уже затем к 3-гидроксиморфинану. Большинство людей не метаболизуют основное количество DXM таким образом, хотя люди с недостатком нормального P450-2D6 будут превращать существенную часть DXM в 3ММ. А 3ММ вероятно не психоактивен, что означает - 5-10% людей с недостатком фермента 2D6 ощутят меньший эффект от DXM (или более точно, не испытают эффектов DXO).
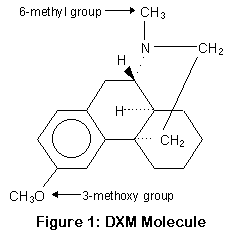
P450-2D6 работает, убирая метокси группу и заменяя ее гидрооксилом (OH) (или точнее, отрезая метил от кислорода); этот процесс называется О-деметиляцией. P450-3A4 и 3A5 заменяют метильную группу водородом (H); это процесс N-деметиляции. Положение метильной и метокси групп показано на рисунке.
9.5.1. Факторы, влияющие на метаболизм DXM
Как говорилось ранее, у некоторых людей отсутствует нормальный фермент P450-2D6. У остальной популяции этот фермент может быть ингибирован несколькими факторами. Многие лекарства ингибируют P450-2D6, особенно флюокситин (ProzacTM). Неполный список лекарств, ингибирующих фермент P450-2D6, дан в Разделе 15.1.
Сам по себе DXM, разумеется, будет конкурировать с другими веществами за P450-2D6 и, что важно, это будет делать и 3-метоксиморфинан (3ММ) (17). Фактически, 3ММ имеет большее сродство к ферменту P450-2D6, чем DXM. Это может объяснить тот факт, что вторая доза DXM "в догонку" производит эффект отличный от первой. Конкуренция за фермент P450-2D6 уменьшит количество DXM, превращающегося в DXO у второй дозы.
|
|
|
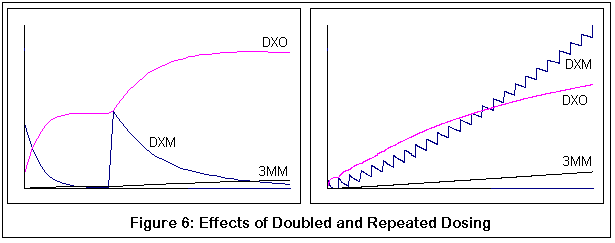
Этот график получен путем компьютерной симуляции метаболизма DXM:
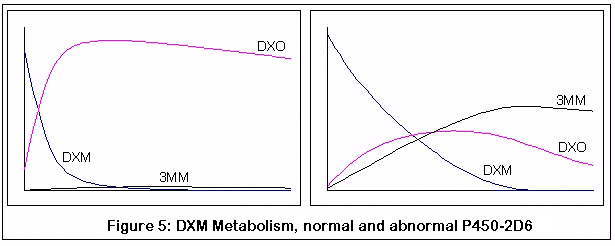
Первая пара графиков представляет метаболизм DXM у нормального индивидуума (слева) и индивидуума без нормального фермента P450-2D6 (справа). Заметьте быстрое и почти полное превращение DXM в DXO у нормального индивидуума, по сравнению с менее эффективным и более медленным превращением у человека с недостатком P450-2D6.
Вторая пара демонстрирует возможные результаты приема дополнительных доз ("в догонку"). Оба графика соответствуют нормальному варианту фермента P450-2D6. Отмечу, что вторая доза DXM не превращается в DXO также быстро (поэтому меньший наклон). Правый график показывает многочисленные дозы, и выгибание вверх метаболитической кривой DXM становится все более явным с каждой дозой.
Между прочим, это все качественные симуляции, а не количественные. Я пытался придерживаться известных значений КМ и VMAX для подходящих реакций [?], но симуляции были дискретным процессом (если честно, мои познания в уравнениях довольно слабы и требуют от меня титанических усилий). Я сравнил мои результаты с обнаруженными скудными данными, сравнения оказались разумными, но, опять же, я могу быть абсолютно неправым. Цель этих графиков - продемонстрировать относительные эффекты изменений в ферментной активности (из-за генетических изменений и конкурентного ингибирования 3-метоксиморфинаном), надеюсь, что этого оказалось достаточно.
У меня нет информации о том, что случается с самим 3-гидроксморфинаном. Он может выделяться напрямую почками, или подлежит дальнейшему метаболизму.
10. Нейрофармакология DXM
10.1. А что такое рецептор? (основы нейрофармакологии)
10.1.1. Структура нервной клетки
У всех клеток существует градиент напряжения между внешней средой и внутренним содержимым клетки; если бы вы измерили это напряжение, то обнаружили бы, что потенциал внутри клетки чуть больше –100 мВ по сравнению с внешней средой. В большинстве клеток этот заряд - следствие переноса ионов в и из клетки, и ни для чего такого не используется. Тем не мене, мускульные и нервные клетки нашли ему применение.
У мускульных и нервных клеток (нейронов) есть "возбудимые" мембраны - заряд мембраны может измениться, а это изменение применяется в специальных целях. В нейронах понижение этого заряда используется для передачи сигнала.
В мозге человека находится 10 миллиардов нервных клеток, и все они делают одну и туже работу: передают сигналы со своих входов (дендритов) на выходы (аксоны). Нейрон немного похож на дерево, только вместо корней и веток у него есть дендриты и аксоны. Грубо говоря, дендриты принимают сигналы, а аксоны - передают (помните, сигнал обозначается изменением потенциала клеточной мембраны).
|
|
|
В конце каждой веточки аксона находится синаптическое утолщение, маленькая, похожая на бутон структура, используемая для посылки сигнала к следующей нервной клетке [не всегда нервной] (об этом позже). Дендриты, наоборот, принимают сигналы и состоят из многих маленьких шишечек, каждая из них принимает сигнал от другого нейрона. Один нейрон может принимать сигналы от тысяч других нейронов.
Некоторые сигналы могут возбуждать мембрану клетки; другие - тормозить возбуждение. Если сумма возбуждающих сигналов минус сумма тормозящих сигналов, полученных дендритами, достаточно велика, то сигнал достигает тела клетки (сомы). Как только достаточно сильный сигнал достигает сомы, он тут же посылается по аксонам и передается дальше.
Это основа индивидуального функционирования нейронов. Тем не менее, это не объясняет, как один нейрон посылает сигнал другому (или, наоборот, как они принимают сигналы). Очень небольшое количество нейронов соединяются друг с другом на физическом уровне, и сигналы просто проходят из клетки в клетку как сигналы от одного транзистора к другому в микросхеме. Однако, большинство нейронов не касаются друг друга; вместо этого, они взаимодействуют с химическими веществами, называемыми нейротрансмиттерами [(нейро)медиаторами] и принимают их с помощью структур, называемых нейрорецепторами.
10.1.2. Нейротрансмиссия
Рецептор - это структура на поверхности или внутри клетки. Если он находится на нервной клетке (нейроне), то часто называется нейрорецептор. Рецепторы необходимы для приема сигналов от определенных химических соединений, и когда они получают эти сигналы, они в основном оказывают влияние на функционирование клетки. Некоторые рецепторы находятся внутри клетки и называются внутриклеточными рецепторами; многие стероидные (тестостерон, эстроген, и т.д.) рецепторы внутриклеточные.
Так как этот раздел о нейрофармакологии, то мы сосредоточимся на нейрорецепторах, обычно называя их рецепторами. Нейрорецепторы располагаются на поверхности (или, реже, внутри) нервной клетки и отвечают на воздействие химических соединений, называющихся нейротрансмиттерами [(нейро)медиаторами]. Некоторые медиаторы воздействуют и на простые клетки; например, ацетилхолин активизирует рецепторы мускульных клеток, заставляя их сокращаться.
Нейромедиаторы подходят к рецептору как ключ к замку, и, в основном, не подходят к любому другому рецептору. Так, у нас есть ацетилхолин - медиатор, и ацетилхолиновые рецепторы. Некоторые рецепторы (которые называются ионные каналы), при активации стимулируют или подавляют мембранный потенциал (тем самым и активность нервной клетки); другие (метаботропные рецепторы) вызывают изменения в характеристиках клетки. Некоторые ионные каналы (кальциевые каналы) делают и то, и другое.
Рецептор на нервной клетке расположен, грубо говоря, таким образом, что может принимать сигналы от другой нервной клетки. И у дендритов есть рецепторы, но рецепторы также могут находиться на теле клетки, на аксонах, или синаптическом утолщении. Сопряжение между двумя нервными клетками называется синапсом.
Нейрорецепторы ионного канала обычно работают очень быстро, и действуют (и выглядят) как ирисовая диафрагма в фотокамере. Нейромедиатор (например, ацетилхолин) связывается со специальным местом на канале, что (из-за электростатических сил) вынуждает канал открыться. Тогда специфичные ионы входят и выходят из нервной клетки, изменяя электрический потенциал. Различные каналы пропускают разные ионы; некоторые ионы (например, натрия) возбуждают нервную клетку, другие (калий и хлор) тормозят ее. Как только медиатор покидает рецептор, канал закрывается, проделав свою работу. Именно эти рецепторы вовлечены в быструю передачу сигналов и проведение импульсов скелетных мышц.
Метаботропные рецепторы играют регулирующую роль. Некоторые из них увеличивают или уменьшают количество рецепторов других типов. Другие вызывают экспрессию [работа гена по производству протеина или образованию фенотипа] генов в клетке. Некоторые (называемые авторецепторами) тормозят выделение совместимого с ними медиатора, такой процесс называется отрицательная обратная связь. Пример системы с отрицательной обратной связью - термостат, чем он горячее, тем слабее его греет печка. В основном, эти рецепторы медленного типа управляют вторичными посредниками [мессенджерами] (которые функционируют как посредники внутри клетки), такими как G-белки. [Собственно говоря, метаботропные рецепторы открывают ионный канал опосредовано, через цепочку биохимических реакций, в частности, посредством активации G-белка].
Любой определенный медиатор может связываться с несколькими различными рецепторами. Например, серотонин (5HT -гидрокситриптамин) активизирует по крайней мере двенадцать подтипов рецепторов (5HT1A, 5HT1B, 5HT1D, 5HT1E, 5HT1F, 5HT2A, 5HT2C, 5HT3, 5HT4, 5HT5, 5HT6, и 5HT7)! Несколько подтипов (вместо всего одного) существуют из-за того, что каждый подтип рецептора вовлечен в разные процессы в нейронах различных типов.
Наркотики действуют на мозг путем изменения нейротрансмиссии [передачи нервного импульса] тем или иным способом. Одни наркотики стимулируют рецепторы, другие блокируюит их; третьи изменяют принцип секреции, распада или переработки медиатора. Наркотики, в большой степени, работают только потому, что влияют на существующие системы передачи нервных импульсов; несмотря на популярное поверие о том, что получение кайфа равносильно поджариванию клеток мозга, на самом деле наркотики создают кайф, просто имитируя, блокируя или по-другому воздействуя на нейротрансмиссию.
Наркотики, имитирующие, блокирующие, или иным способом воздействующие на работу определенного медиатора, не одинаково действуют на рецепторы всех подтипов. Например, LSD действует на рецепторы 5HT2A и 5HT2C; буспирон - на 5HT1A. Соответственно, они оказывают различные эффекты; LSD - психоделик, а буспирон - успокаивающее средство.
Различные вещества могут привязываться к одному и тому же рецептору, в то же время по-разному влияя на него. Агонист - связывающееся с рецептором и активирующее его вещество. Частичный агонист - агонист не полностью активизирующий рецептор. Антагонист связывается с рецептором и прекращает его функционирование.
Одна из интесных особенностей частичных агонистов - это их способность "нормализовать" уровень активности рецептора. В присутствии малого количества медиатора, частичный агонист увеличивает функционирование рецептора. Тем не менее, в присутствии большого количества медиатора, частичный агонист ограничивает активность рецептора; фактически, многие антагонисты на самом деле могут являться частичными агонистами. До сих пор обсуждается вопрос - является ли LSD антагонистом или частичным агонистом рецептора 5HT2C.
Антагонисты могут связываться с тем же местом, что и медиаторы, таким образом "конкурируя" с медиатором - они называются конкурентными антагонистами. Или они могут связываться с другим местом на рецепторном комплексе, и теперь, если медиатор и достигнет места связывания, рецептор не активизируется. Такие антагонисты называются неконкурентными. Заметьте, что в обоих случаях, наркотик связывается временно; при постоянном связывании (эффективном разрушении рецептора) это будет необратимым антагонизмом.
Вот в чем заключается важное различие между конкурентным и не конкурентным антагонистом: при блокаде рецепторов конкурентным антагонистом, они все еще могут быть активизированы достаточным количеством медиатора; При блокаде неконкурентным антагонистом никакое сколь угодно большое количество медиатора не сможет активизировать рецепторы (до тех пор, когда неконкурентный агонист уйдет).
Довольно причудливая аналогия может быть проведена между работой медиатора и туалетом. В этом случае, туалет - это рецептор, вы - медиатор, активизирующий туалет, нажимая ручку слива. Если ваш маленький брат придет и спустит воду за вас - он агонист. Если он на время задержит ручку на полпути - он частичный антагонист. Если он не даст вам нажать на ручку - он конкурентный антагонист. Если он устроит засор из туалетной бумаги - он неконкурентный антагонист, а если он оторвет ручку - он необратимый антагонист.
Медиаторы, принадлежащие к группе биогенных аминов, включают в себя ацетилхолин, норадреналин, дофамин, серотонин (5HT), и гистамин. Они происходят из аминокислот (холина, тирозина, триптофана и гистидина соответственно), и в основном, играют регулирующую роль, и являются обычными мишенями рекреационных наркотиков. Например, LSD, ДМТ и псилоцибин направлены на 5HT рецепторы; амфетамины вызывают вырабатывание дофамина и норадреналина; кокаин блокирует обратное всасывание дофамина (таким образом, продлевая его активность); МДМА вызывает вырабатывание 5HT и дофамина; и т.д. Наиболее полный список рекреационных наркотиков и их воздействие на нейрорецепторы даны в Разделе 15.2.
Нейропептидные медиаторы включают в себя огромное количество пептидов (цепочек аминокислот) таких, как нейропептид Y, ангиотензин, эндорфины, вещество P, и так далее. Единственные рекреационные наркотики, нацеленные на нейропептидные рецепторы - опиаты. Они связываются с мю, каппа и дельта опиоидными рецепторами. Опиоидные рецепторы (очевидно) вовлечены в болевое и поведенческое подкрепление. Вазрпрессин, ноотроп (Smart drug) - также нейропептидный медиатор.
Аминокислотные медиаторы включают в себя ГАМК (гамма-аминомасляная кислота), глутамат, и аспартат. Их рецепторы - два вида ГАМК рецепторов, NMDA рецепторы, AMPA (бывший квисгулат), рецепторы каината и метаботропные рецепторы (они все отвечают на воздействие глутамата и аспартата). На ГАМК рецепторы действуют бензодиазепины типа диазепама (ValiumTM), барбитураты, алкоголь; NMDA рецепторы подвержены влиянию PCP, кетамина, алкоголя и DXM.
А еще есть такие рецепторы, которые не входят ни в один класс. Недавно определен анандаминовый рецептор - цель ТКГ (THC) - действующего вещества марихуаны. Аденозиновый рецептор, имеющий тенденцию тормозить нервную активность, блокируется кофеином (в этом и есть его стимулирующий эффект). Сигма рецептор, ранее отнесенный к опиоидным рецепторам, в настоящее время считается отдельным. Гамма-гидроксибутират, GHB, видимо тоже нацеливается на специфический рецептор.
У каждого рецептора может более одного места связывания (место, к которому вещество может прикрепляться, в основном воздействуя на активность рецептора (иногда называется "сайт"). Например, у NMDA комплекса канал/рецептор их семь (глутамат, глицин, ион магния, цинка, сайт открытого канала PCP, сайт полиамина и сайт фосфорилирования). У большинства их меньше; NMDA-канал - очень сложный рецептор.
Вольт-зависимые ионные каналы аналогичны быстродействующим рецепторам с ирисовым затвором, с тем исключением, что открываются разностью потенциалов клеточной мембраны. Они обычно передают сигналы по нервным волокнам, или вынуждают окончание аксона выделить медиатор. Натрий, калий, кальций и хлор (Na+, K+, Ca2+, и Cl-) - это те самые ионы, которые проводят эти каналы. Тетродотоксин, активный ингредиент в "порошке зомби" [выделяется из рыбы-собаки (фугу)], блокирует натриевые каналы. NMDA рецепторы обладают некоторыми чертами вольтзависимых ионных каналов (см. ниже).
10.2. Что такое Сигма-рецепторы?
Открытые в 1976 году, сигма рецепторы (сигма часто пишется по-гречески σ) в настоящее время одна из самых смущающих вещей в нейрофармакологии. Наши знания о сигма рецепторах блекнут перед нашим невежеством; фактически, то, что мы знаем абсолютно точно (или думаем, что абсолютно точно), может быть сведено в следующий абзац:
По всему мозгу и телу рассыпаны места (сигма-сайты связывания), где группа химических соединений (сигма-лигандов) обычно застревает. Мы не знаем, находятся ли они внутри или снаружи клеток. Мы не знаем, изменяет ли что-либо прилипание к ним сигма-лигандов, кроме как в семявыносящем протоке. Мы на самом деле не знаем, что они делают, если хоть что-то. Мы не знаем, для чего они нужны, почему они тут, использует ли их тело. Они могут быть нейрорецепторами, стероидными рецепторами, рецепторами внутриклеточных посредников, регуляторами роста, ферментами или чем-то совершенно иным.
Другими словами - приготовьтесь к непоняткам. Не беспокойтесь - все такие.
Сигма-рецепторы сначала предполагались опиоидными, так как многие производные морфина связываются там. Тем не менее, эта классификация, вероятно, ложная, и эндогенные опиоиды проявляют малую сигма активность. Большинство характеристик опиатов передаются мю (μ), каппа (κ) и дельта (δ) рецепторами. Существуют, по крайней мере, два сигма рецептора; третий (сигма3, соответственно) был открыт недавно.
Некоторые исследователи предполагали, что сигма рецепторы - вовсе не рецепторы, а просто места связывания ферментов. С другой стороны, сигма-лиганды воздействуют на мускулы семявыводящих протоков гвинейской свиньи, чего не случилось бы, если бы сигма рецепторы не были рецепторами. Сигма рецепторы могут быть предназначены для гормонов или внутриклеточных посредников [мессенджеров], а не медиаторов, так как они находятся по большому счету в микросоме, а не на поверхности клетки.
10.2.1. Сигма1 рецепторы и общая информация
Большинство из известного о сигма рецепторах, видимо, относится к сигма1, чем к сигма2 рецептору (хотя это ни в коей мере не универсальное правило). Я соотношу следующую информацию с сигма1 рецепторами, но не принимайте это как непререкаемый факт. Я ожидаю, что многое окажется неправильным. К счастью, долго ждать не придется; исследования в области сигма рецепторов быстро прогрессируют.
10.2.1.1. Эндогенные лиганды
Медиаторов для сигма1 рецепторов не обнаружено, хотя есть некоторые предположения. Обычное название для (неопределенного) сигма1 медиатора - "эндопсихозин", раньше известный как "энджелдастин" [от Angel Dust - одно из названий PCP]. Прогестерон нацелен на сигма1 рецепторы в плаценте, он и другие стероидные гормоны могут быть природными лигандами для сигма1 рецепторов. Если это верно, то, возможно, что некоторые воздействия половых гормонов на мозг передаются сигма1 рецептором. Предполагалось, что субстанция P (пептидный нейромедиатор) является эндогенным сигма1 лигандом, но эта гипотеза была отвергнута. Дегидроэпиандростерон (DHEA) может быть сигма1 агонистом, а прогестерон антагонистом.
10.2.1.2. Местоположение и функции в мозге
Наиболее плотно сигма рецепторы располагаются в коре головного мозга (281), nucleus accumbens, и в коре, а также менее плотно в лимбических областях и экстрапирамидальной моторной системе. Это представляет интерес из-за странных эффектов DXM, оказываемых на движение, которые могут иметь отношение к сигма активности в коре головного мозга и экстрапирамидальной моторной системе.
Сигма1 (и возможно сигма2) рецепторы функционально соединены с некоторыми другими рецепторами; стоит отметить никотиновые (никотинподобные) ацетилхолиновые рецепторы и NMDA рецепторы. Они действительно могут находиться на или рядом с NMDA рецепторами.
Связь с никотиновыми рецепторами может быть прямой - сигма активация вызывает изменение в работе никотиновых рецепторов. Я не знаю, повлияет ли изменение состояния никотиновых рецепторов на эффекты, производимые никотином на мозг; некоторые люди сообщали, что табак вызывает сильные реакции во время использования DXM.
Сигма агонисты (и/или, возможно, антагонисты), вероятно, воздействуют на работу памяти, обращая нарушение памяти, вызванное такими наркотиками как п-хлорамфетамин и МК-801 (наркотик, аналогичный кетамину). DTG, (+)-пентазоцин (pentazocine), и SKF-10047 - все улучшают нарушение памяти, вызванное МК-801. С другой стороны, NE-100 - предполагаемый сигма антагонист, вероятно, помогает при нарушении памяти, вызванном NMDA антагонистами. DTG - сигма агонист - обращает нарушение памяти, вызванное угарным газом.
Многие средства - сигма агонисты и антагонисты - на самом деле могут быть частичными агонистами. Другая возможность - оптимальный уровень сигма активности это здоровая середина; одно исследование выявило колоколообразную зависимость активности от дозы сигма агонистов. Это аналогично эффекту многих ноотропов, особенно холинергиков - прием большого количества может быть хуже отказа от приема. Эта аналогия может служить основой для связи сигма и ацетилхолиновых рецепторов.
И сигма1 агонисты, и антагонисты могут защитить NMDA рецепторы от глутаматной токсичности. Одно исследование обнаружило, что сигма антагонисты защищают гиппокампальные клетки от гипоксии и гипогликемии, а это тоже может быть связано с NMDA рецепторами. Морфин обладает непрямым воздействием на NMDA рецепторы, которое передается вероятно сигма рецепторами, возможно сигма1. Вероятно, все эти эффекты передаются через никотиновые рецепторы, то есть сигма1 могут не на прямую контролировать работу NMDA рецепторов.
10.2.1.3. Поведенческие эффекты
Поведенческие эффекты сигма1 рецепторов полностью не установлены. Тем не менее, сигма1 (и сигма2) рецепторы, по-видимому, воздействуют на моторные функции, производя и усиливая локомоцию. Часть этих эффектов может происходить в мозжечке; может играть роль и выброс дофамина. Возможно, это и является источником любопытных эффектов DXM на движение, походку, включая походку моряка и Робо-шарканье.
|
|
|
