 |


Поглощение энергии света листьями
|
|
|
|
Лист как орган растения, приспособленный к фотосинтезу, сформировался в результате длительного эволюционного процесса. Он представляет собой эффективную систему для поглощения и преобразования энергии света в ходе фотосинтеза. Структура листа обеспечивает наиболее полное поглощение квантов света, поступление углекислого газа из атмосферы к хлоропластам, а также возможность оттока ассимилятов из автотрофных клеток.
Как все физические тела, лист поглощает, пропускает и отражает падающие на него солнечные лучи. Лучистая энергия, посылаемая от Солнца к Земле, представляет собой электромагнитные колебания с разными длинами волн. Около 40–45 % этой энергии приходится на область от 380 до 720 нм. Эта часть спектра воспринимается как видимый свет. Здесь располагаются известные цвета радуги: фиолетовый, синий, голубой, зеленый, желтый, оранжевый и красный. К этой области со стороны более коротких волн примыкает ультрафиолетовая радиация, а со стороны более длинных – инфракрасная радиация. Последняя воспринимается как тепло. Инфракрасные лучи не участвуют непосредственно в фотосинтезе, но регулируют другие процессы жизнедеятельности. Коротковолновая радиация (ультрафиолетовые, γ-лучи и космические лучи), по-видимому, играет огромную роль в мутагенезе растений, в изменении их наследственности.

Участок видимого спектра, поглощаемый пигментами хлоропластов (380 – 700 нм), получил название фотосинтетически активной радиации (ФАР).

Лист поглощает 85 % видимого света, пропускает 5 % и отражает 10 %. Инфракрасная радиация поглощается на 25 %, пропускается на 30 % и отражается на 45 %.
Адаптация растений к определенным экологическим условиям мало влияет на поглощение, пропускание и отражение света. Средние величины, характеризующие оптические свойства листа у разных жизненных форм, тоже отличаются мало. Исключения составляют водные растения и растения сухих мест: у подводных листьев водных растений значительно увеличивается пропускание, у суккулентов, напротив, оно приближается к нулю.
|
|
|
Оптические свойства листа могут изменяться. При недостатке воды и большой освещенности восковой слой и опушенность листовой пластинки увеличиваются. Волоски увеличивают отражение света. В тени плохо развивается столбчатая паренхима, что ухудшает поглощение света. Как уже было сказано, количество хлоропластов в листе зависит от условий азотного питания и водоснабжения.
Набухание хлоропластов увеличивает рассеивание ими света. Это показывает важность для фотосинтеза водного гомеостаза клетки. Поглощение света регулируется и движением хлоропластов в клетке. В условиях недостаточного освещения хлоропласты располагаются у клеточных стенок, перпендикулярных солнечным лучам, а в условиях излишней освещенности – у клеточных стенок, параллельных им. Первое расположение увеличивает, а второе уменьшает поглощение света.
Листовая поверхность, поглощающая свет, достигает значительных размеров. Так, общая листовая поверхность у растений умеренной зоны, растущих на 1 га пашни, составляет примерно 5 га, а в южных широтах с влажным климатом – 10–15 га. Размещение листьев в пространстве позволяет растениям использовать как прямой, так и рассеянный свет, падающий под разными углами. Здесь следует вспомнить основной закон фотохимии – закон Гротгуса: фотохимический процесс, следовательно, и фотосинтез, совершаются лишь под действием тех лучей, которые поглощаются. Однако растения умеренной зоны используют для фотосинтеза 1–2% (максимум 5 %), а тропические – 5–6 % и даже до 15 % поглощенного видимого света. Остальная энергия расходуется на испарение воды. Свет поглощается пигментами листа.
|
|
|
Коэффициент полезного действия фотосинтеза листа показывает, сколько процентов поглощенной световой энергии запасено в форме энергии химических связей при превращении диоксида углерода в углеводы. В процессе фотосинтеза на 1 г полученного углевода используется 25,9 кДж энергии. Умножение этого коэффициента на интенсивность брутто-фотосинтеза дает количество связанной энергии (величины пересчитываются на одинаковую поверхность и одинаковый промежуток времени). КПД фотосинтеза обычно выражают по отношению либо к падающей, либо к поглощенной растениями ФАР. В большинстве случаев листья работают с КПД меньше 5–10 %, но у некоторых видов в особо благоприятных условиях он достигает 15 % (у С4-злаков 24 %).
Несмотря на высокую эффективность начальных фотофизических и фотохимических стадий (около 95 %), в урожай переходит лишь несколько процентов солнечной энергии. Потери обусловлены лимитированием процесса на биохимическом и физиологическом уровнях, а также неполным поглощением света. Во всех сообществах получаемая солнечная радиация используется неэффективно. К факторам, ограничивающим первичную продукцию на суше, относятся: 1) недостаток воды, ограничивающий скорость фотосинтеза; 2) нехватка элементов минерального питания, замедляющая скорость образования ассимилирующей ткани и снижающая эффективность фотосинтеза; 3) неблагоприятная для роста температура; 4) попадание большой части радиации мимо фотосинтезирующих органов (из-за сезонного опадения листьев, действия фитофагов и паразитов); 5) низкая эффективность фотосинтеза в листьях (даже при идеальных условиях в наиболее продуктивных сельскохозяйственных системах она редко бывает выше 10 % ФАР). При изменяющихся условиях ассимиляции КПД растительных сообществ суши в среднем ниже 1 – 3%. Максимальная эффективность фотосинтеза интенсивных зерновых культур при идеальных условиях 3 –10 %. А если рассматривать планету в целом, то КПД падающей ФАР составляет всего около 0,2.
Строение хлоропластов
Хлоропласт высшего растения похож на линзу, диаметр которой 5–8 мкм, а толщина – около 1 мкм. Внутренняя мембрана образует выросты, параллельные длинной оси органеллы и получившие название тилакоидов стромы. Они соединяют между собой граны, тоже состоящие из тилакоидов. Тилакоид (от греч. thylacoides – мешковидный) представляет собой плоский мешочек, ограниченный однослойной мембраной. В тилакоидах находятся пигменты и ферменты, принимающие участие в световой фазе фотосинтеза. В одной гране может находиться 10–30 тилакоидов, а в хлоропласте – до 100–150 гран. Все пространство между гранами заполнено бесцветным раствором – стромой. В строме содержатся ферменты, участвующие в восстановлении СО2.
|
|
|
Хлоропласты, имеющие такое строение, получили название гранальных. Однако у ряда растений (кукуруза, сахарный тростник, просо и др.) есть еще хлоропласты без гран, называемые аграналъными.
В строении хлоропласта мы можем наблюдать один из принципов построения живой материи – иерархию: так же, как организм состоит из органов, орган – из клеток, а клетка – из органелл, хлоропласт состоит из стромы и тилакоидов, тилакоиды образуются из мембран и сами могут образовывать граны.
В хлоропластах находятся все фотосинтетические пигменты (5–10 % от его сухого вещества). Кроме того, хлоропласт содержит 35–55 % белков, 30–50 % липидов, 2–3 % РНК, 0,5 % ДНК, а также 5 % золы, основными компонентами которой являются калий, фосфор – 50 %, железо – 80 %, медь – 50 %, магний, цинк – 70 % от всего содержащегося в листе количества. В хлоропластах имеются своя специфическая кольцевая ДНК, которая отличается от ядерной и передается по наследству через пропластиды яйцеклетки, а также РНК, рибосомы и ферменты, необходимые для синтеза белка.
Гранальные хлоропласты могут делиться, но чаще они образуются из пропластид. Пропластида окружена двойной мембраной. Из внутренней мембраны пропластиды сначала возникает проламеллярное тело. Затем, если света достаточно, из него формируются тилакоиды и граны. В мембраны тилакоидов встраиваются вновь синтезированные молекулы пигментов. Одновременно с ростом клеток растут и хлоропласты. Предполагают, что агранальные хлоропласты возникают в процессе формирования листовой пластинки из гранальных, так как сначала они тоже имеют граны. В клетке в среднем находится до 50 хлоропластов. В старой клетке хлоропласты разрушаются. Хлоропласты могут двигаться вместе с цитозолем (пассивно) и самостоятельно.
|
|
|
Пигменты фотосинтеза
Пигменты – это вещества, избирательно поглощающие свет в видимой части спектра. При освещении белым светом их окраска определяется теми лучами, которые они пропускают или отражают. Если вещество не поглощает свет, то к нам в глаз приходят все лучи видимого спектра, и для нас это вещество белое. Если вещество поглощает все лучи видимого спектра, то для нас оно черное.
Способность пигментов поглощать свет связана с наличием в их молекулах правильно чередующихся двойных и одинарных связей. Это так называемые сопряженные двойные связи. Между двумя атомами, связанными двойной связью, находится 4 электрона. Если система состоит из сопряженных связей, то половина этих π-электронов может свободно перемещаться вдоль всей системы. Поглотив квант света, такой электрон способен оторваться от молекулы пигмента, т.е. пигмент становится донором электронов для восстановления вещества.
Пигменты, участвующие в фотосинтезе высших растений, делятся на 2 группы: хлорофиллы – зеленые пигменты – и каротиноиды – желтые. Есть два хлорофилла – хлорофилл а и хлорофилл b. Хлорофилл а – сине-зеленый, а хлорофилл b – желто-зеленый. Хлорофиллы не растворяются в воде, но хорошо растворимы в органических растворителях и легко изменяются под действием кислот, солей и щелочей. В листе содержится примерно в три раза больше хлорофилла а по сравнению с хлорофиллом b.
По химической природе хлорофилл представляет собой сложный эфир дикарбоновой кислоты – хлорофиллина – и двух спиртов: метанола (СН3ОН) и фитола (С20Н39ОН):
В основе молекулы хлорофилла лежит порфирин, состоящий из четырех пиррольных колец, соединенных метиновыми мостиками (—СН=). Хлорофилл относится к тетрапирролам. Именно 4 пиррольных кольца и метиновые мостики создают сопряженные связи. В центре молекулы хлорофилла расположен атом магния, который соединен с четырьмя атомами азота пиррольных колец. Следовательно, хлорофилл относится к магний-порфиринам.
Хлорофилл b отличается от хлорофилла а тем, что у него ко второму пиррольному кольцу присоединена не метальная, а альдегидная группа, поэтому хлорофилл b содержит кислорода на один атом больше, а водорода – на два атома меньше.

| 
| 
|
|
|
|
Благодаря наличию сопряженных двойных связей с подвижными π-электронами и атомов азота с неподеленными электронами молекула хлорофилла в интактной (неповрежденной) клетке способна к обратимым окислительно-восстановительным реакциям под действием света. Азот пиррольных колец может окисляться (отдавать электрон) или восстанавливаться (присоединять электрон).
Молекулу хлорофилла делят на две части: порфириновое ядро и фитолъный хвост. Фитольный хвост в 2 раза длиннее. Благодаря атомам кислорода, азота и магния порфириновое ядро гидрофильно. Фитольный хвост – это углеводородная часть, следовательно, он гидрофобен.
Таким образом, молекула хлорофилла полярна. Эта полярность молекулы обусловливает ее расположение в мембранах хлоропласта: фитольный хвост располагается в гидрофобной части мембраны тилакоида, а порфириновое ядро – в гидрофильной. Имея разные свойства, обе части молекулы хлорофилла выполняют разные функции: порфириновое ядро поглощает свет, а фитольный хвост играет роль якоря, удерживающего молекулу хлорофилла в определенной части мембраны тилакоида.
Полученный из листа хлорофилл легко реагирует с кислотами и щелочами. При взаимодействии со щелочью образуются два спирта – метанол и фитол – и щелочная соль хлорофиллина.
Щелочь «отрезает» от молекулы хлорофилла фитольный хвост, в результате образующаяся соль теряет способность растворяться в бензине, но сохраняет зеленый цвет. Следовательно, растворимость хлорофилла в бензине, его гидрофобность обусловлены остатком фитола, а поглощение света связано с порфириновым ядром.
При действии слабой кислоты хлорофилл теряет зеленый цвет, образуется красно-бурое вещество феофитин, у которого атом магния замещен на два атома водорода.
Следовательно, атом магния тоже влияет на поглощение света хлорофиллом. В настоящее время роль магния в поглощении света связывают с тем, что с его помощью четыре пиррольных кольца располагаются в одной плоскости. Если молекула по какой-то причине приобретает другую форму, то взаимодействие π-электронных облаков нарушается, цепь сопряжения разобщается, цвет пигмента изменяется или исчезает. С помощью магния молекулы хлорофил
ла соединяются с другими молекулами этого пигмента. Кроме того, магний нужен для сохранения молекулой хлорофилла своего возбужденного состояния.
В естественных условиях образование феофитина происходит при старении листьев, осенью, под влиянием неблагоприятных факторов. В результате листья желтеют. В природе появление феофитина вызвано увеличением проницаемости мембран и проникновением в хлоропласт кислого клеточного сока. Поскольку избирательная проницаемость мембран увеличивается под действием любого фактора, то и листья желтеют под действием низких и высоких температур, дефицита воды и ее избытка. Этот факт лишний раз доказывает важность этого свойства мембран.
Кроме пиррольных колец, в состав молекулы хлорофилла входит еще карбоциклическое кольцо с высокоактивной кетогруппой. Предполагают, что эта группа участвует в окислении воды.
Каротиноиды – полиеновые углеводороды красного, желтого и оранжевого цветов, производные изопрена, содержащие 40 атомов углерода. Каротиноиды представляют собой цепи, обладающие, как и хлорофиллы, сопряженными двойными связями. На обоих концах цепи находятся иононовые кольца. Каротиноиды присутствуют в хлоропластах всех растений. Они входят также в состав хромопластов. Свое название эти вещества получили от латинского названия моркови Daucus carota, в корнеплоде которой они содержатся в большом количестве. В зеленых листьях каротиноиды обычно незаметны из-за присутствия хлорофилла, но осенью, когда хлорофилл исчезает, окрашивают листья в желтый и оранжевый цвета.
Каротиноиды делятся на 2 группы: каротины (С40Н56) и ксантофиллы (C40H56O2 и С40Н56О4). Каротины представляют собой углеводороды (тетратерпены), а ксантофиллы – содержат дополнительные гидроокси- и эпоксигруппы. У высших растений известны два каротина (α-каротин и β-каротин) и 4 ксантофилла (лютеин, виолаксантин, зеаксантин и антероксантин).
β-Каротин имеет два β-иононовых кольца (двойная связь между С5 и С6 атомами). α-Каротин отличается от β-каротина тем, что у него одно кольцо β-иононовое, а второе – е-иононовое (двойная связь между С4 и С5 атомами).
Лютеин – производное α-каротина, а зеаксантин – β-каротина. Эти ксантофиллы имеют по одной гидроксильной группе в каждом иононовом кольце. Виолаксантин имеет еще дополнительно 2 атома кислорода по двойным связям С5 и С6. Каротиноиды нерастворимы в воде, но хорошо растворяются в бензоле, бензине, ацетоне, сероуглероде.


Все пигменты поглощают свет избирательно. Так, если пропустить белый свет через раствор хлорофилла, а затем разложить его с помощью призмы, то можно увидеть, что отдельные лучи спектра окажутся сильно поглощенными и на их месте будут черные полосы. Другие лучи будут лучше проходить через раствор. В результате мы получим так называемый спектр поглощения хлорофилла.

Хлорофиллы поглощают максимально красные и сине-фиолетовые лучи, хуже поглощают оранжевые, желтые и голубые, почти не поглощают зеленые и отражают дальние красные лучи. Небольшие различия в строении молекул хлорофилла а и b обусловливают некоторые различия в поглощении ими света. У хлорофилла b полоса поглощения в красной части спектра сдвинута в сторону коротковолновых лучей, а в сине-фиолетовой части – в сторону длинноволновых лучей.
Желтые пигменты поглощают лучше те лучи, которые плохо поглощают зеленые пигменты. Максимально они поглощают сине-фиолетовые, хуже – голубые, немного поглощают зеленые лучи, не поглощают красные, желтые и оранжевые. Почему разные пигменты поглощают разный свет? Это зависит от числа и расположения двойных связей, присутствия в них ароматических колец и атома металла в молекуле пигмента.
Хлорофилл может флуоресцировать. Флуоресценция – явление свечения некоторых веществ при их освещении. Если источник света убрать, то флуоресценция прекратится. Испускаемый при флуоресценции свет всегда имеет большую длину волны, чем поглощенный, так как часть поглощенной энергии превращается в тепловую. Хлорофилл флуоресцирует красным (вишневым) светом. Флуоресценция доказывает, что не вся поглощенная хлорофиллом энергия используется на фотосинтез. Флуоресценция тем сильнее, чем меньше поглощенной энергии света используется для фотосинтеза.
Изучение спектров поглощения показало, что хлорофилл а в живом листе существует в нескольких формах, химически одинаковых, но различающихся по поглощению света. Это обусловлено разными причинами. Во-первых, молекулы хлорофилла при близком расположении могут соединяться друг с другом в агрегаты. Во-вторых, они взаимодействуют с белками и липидами мембран тилакоидов. Связь между молекулами белка и хлорофиллом осуществляется путем образования нестойких комплексов, образующихся при взаимодействии кислотных групп белковых молекул и азота пиррольных колец. В-третьих, это связано с динамическим состоянием молекул хлорофилла в мембранах: они непрерывно разрушаются под действием света (фотодеструкция), вместо них в мембрану встраиваются новые молекулы.
Роль пигментов в фотосинтезе. Из физики известно, что только поглощенный свет может производить химическое действие; поглощенный квант света активирует лишь одну молекулу пигмента; вся энергия кванта поглощается только одним электроном, который в результате поднимается на более высокий энергетический уровень; на основном уровне остается «электронная дырка» (электронная недостаточность). Такое состояние молекулы пигмента называют возбужденным; при этом поглощенная энергия запасается в виде энергии электронного возбуждения. В тексте или на рисунке около названия или формулы возбужденной молекулы ставят звездочку (*).
Электрон может находиться на верхнем энергетическом уровне очень недолго: 10-10—10-12 с. Затем он теряет поглощенную энергию и возвращается на свое место или переходит к другому веществу, при этом поглощенная энергия может выделиться в виде света, тепла или использоваться для синтеза АТФ из АДФ. В последнем случае световая энергия превратилась в химическую.
Зеленые и желтые пигменты играют в фотосинтезе разную роль. Еще в 1932г. Р. Эмерсон (США) разделил фотосинтетические пигменты на две группы: пигменты-сборщики света и пигменты-ловушки. Пигменты-сборщики – это пигменты, поглощающие свет и передающие поглощенную энергию квантов пигменту-ловушке. Пигмент-ловушка – пигмент, который, получив энергию, может потерять электрон, что приводит к разделению зарядов.

В результате молекула пигмента-ловушки окисляется. Разделение зарядов является функцией реакционного центра. Реакционный центр представляет собой пигмент-белковый комплекс, содержащий пигмент-ловушку.
У высших растений выделено два основных реакционных центра, включающих пигменты П700 или П680. П700 (Р700) и П680 (Р68о) – это пигменты-ловушки, на долю которых приходится около 1 % всех фотосинтетических пигментов. Они представляют собой особые формы хлорофилла а. П (Р) – это первая буква русского (или латинского) слова «пигмент» (pigmentum), а цифра около буквы показывает, что максимум поглощения приходится у пигмента на длину волны 700 или 680 нм. Им принадлежит главная роль в фотосинтезе. В результате первичного разделения зарядов в реакционных центрах энергия возбужденного состояния хлорофилла преобразуется в химическую энергию.

Остальные пигменты (99 %) являются сборщиками; это – вспомогательные пигменты, образующие светособирающие (антенные) комплексы (ССК). Каждый ССК содержит несколько сотен молекул хлорофиллов и каротиноидов. Пигменты-сборщики передают поглощенную ими энергию с помощью резонанса пигменту-ловушке. Передача энергии происходит от каротиноидов к хлорофиллу и от одной молекулы хлорофилла к другой; от пигментов, поглощающих свет меньшей длины волны, к пигментам, поглощающим свет большей длины, т.е. от коротковолновых пигментов к более длинноволновым.
Время передачи энергии от одной молекулы хлорофилла к другой равняется 1-2·10-12 с, от молекулы каротиноидов к хлорофиллу – 4 · 10-10с.
Для того чтобы энергия возбуждения молекулы одного пигмента могла передаться молекуле другого, они должны располагаться в определенной последовательности и между ними должны быть очень маленькие расстояния (1,15 нм). Это возможно благодаря тому, что пигменты-сборщики располагаются в мембранах тилэкоидов. В светособирающем комплексе молекулы пигментов ориентированы определенным образом относительно мембраны хлоропласта и друг друга. Пигменты-сборщики, передавая поглощенную энергию ловушке, способствуют использованию для фотосинтеза тех лучей, которые самим пигментом-ловушкой не поглощаются.
В процессе передачи часть энергии теряется в виде тепла. Потеря энергии приводит к превращению квантов в более мелкие, т.е. с большей длиной волны. Именно поэтому пигменты-ловушки П700 и П680 являются более длинноволновыми. Перенос энергии в обратном направлении невозможен.
Итак, функция пигментов ССК – поглощение и передача энергии квантов в реакционные центры на П680 и П700, которые осуществляют фотохимические реакции. Фотохимические процессы в реакционных центрах приводят к быстрому запасанию энергии квантов света в форме лабильных соединений с высоким энергетическим потенциалом. Дальнейшие реакции фотосинтеза направлены на преобразование энергии света в более стабильную форму восстановленного НАДФ и АТФ, которые затем используются для синтеза углеводов и других стабильных органических соединений (темновая фаза фотосинтеза).
Каротиноиды не только передают поглощенную световую энергию хлорофиллу, но и выполняют защитную функцию. Поглотившая квант света молекула хлорофилла способна прореагировать с кислородом атмосферы, в результате чего молекула хлорофилла переходит в основное состояние и образуется активный кислород, который может вызвать повреждение хлорофиллов и мембран. Каротиноиды защищают клетку от образования активного кислорода: забирают энергию от хлорофилла и выделяют ее в виде тепла. У растений-мутантов, не имеющих каротиноидов, в аэробных условиях хлорофилл быстро разрушается, и они погибают.
Каротиноиды защищают фотосинтетический аппарат и от слишком высокой интенсивности света, которая может его повредить. Механизм защиты связан с превращением виолаксантина в зеаксантин; в темноте происходит обратный процесс.
Зависимость образования пигментов от внутреннего состояния растений и внешних условий. Первые признаки зеленения выращенных в темноте этиолированных растений наблюдаются через 2-4 ч после начала освещения. Оптимальным для синтеза хлорофилла является освещение 200 Вт/м2. При сильном повышении интенсивности света биосинтез хлорофилла начинает замедляться. Поэтому световые листья имеют меньшую концентрацию хлорофилла, чем темновые (эффект Шталя).
В опытах В.Н. Любименко показан нижний предел освещенности для образования хлорофилла. У проростков ячменя и овса таким пределом было освещение лампой накаливания мощностью 10 Вт. При исследованиях было выявлено, что образование хлорофилла идет интенсивнее на прерывистом свете.
Образование хлорофилла зависит от температуры. Оптимальной температурой для накопления хлорофилла является 20- 25 °С, причем особое значение температура имеет для темновых реакций синтеза предшественников хлорофилла. Поэтому растения ранней весной часто не зеленеют и остаются этиолированными до тех пор, пока температура не превысит 10 °С. На скорость образования хлорофилла и особенно его предшественников оказывает влияние оводненность тканей. При недостаточной водообеспеченности биосинтез хлорофилла затормаживается и наблюдается его распад. Пожелтение листьев при сильных засухах - обычное внешнее проявление водного дефицита.
Важное значение для образования хлорофилла имеют условия минерального питания. Растениям прежде всего необходимо хорошее обеспечение азотом и магнием, которые входят в состав хлорофилла. При недостатке этих элементов наблюдается хлороз - утрата зеленой окраски листьев. Причиной хлороза может быть и дефицит железа. Железо входит в состав ферментов, катализирующих важнейшие звенья биосинтеза хлорофилла. При недостатке меди хлорофилл легко разрушается, так как нарушаются комплексы между хлорофиллом и соответствующими белками.
На общий ход биосинтеза хлорофилла влияет возраст листьев и растения в целом. В молодых листьях биосинтез хлорофилла идет в 10-15 раз быстрее, чем в старых. Молекулы хлорофилла не могут служить неограниченно долго. Часть их постепенно разрушается, заменяясь вновь синтезированными. «Молодые» молекулы хлорофилла в химическом отношении тождественны «старым», но имеют иную связь с белками и работают более активно. Таким образом, биосинтез и распад хлорофилла представляют сложный комплекс биохимических реакций, находящихся под контролем как генетических факторов, так и условий произрастания растений.
Тема 3 Этапы фотосинтеза
1. Световая фаза фотосинтеза
2. Фотосинтетическое фосфорилирование
3. Пути фиксации СО2 при фотосинтезе
4. Фотодыхание
Световая фаза фотосинтеза
Традиционно в процессе фотосинтеза различают две стадии: световую и темновую. Световая стадия происходит только в присутствии света в мембранах тилакоидов. Темновая стадия – не совсем удачное название. Если мы говорим, что световая фаза названа так, чтобы подчеркнуть зависимость всех протекающих в ней реакций от света, то название «темновая фаза» означает, что все реакции, в ней протекающие, от света не зависят и проходят в темноте. Но это не совсем точно. Очень многие реакции темновой фазы фотосинтеза зависят от света, так как ферменты, катализирующие эти реакции, индуцируются светом. Поэтому, согласно современным представлениям, выделяют три фазы фотосинтеза: фотофизическую, фотохимическую и химическую (синтетическую).
В фотофизической фазе происходит поглощение квантов света пигментами, их переход в возбужденное состояние и передача энергии к другим молекулам фотосистемы.
В фотохимической фазе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электрон-транспортной цепи, что заканчивается синтезом АТФ и НАДФН.
Химическая (синтетической) фаза происходит без участия света и включает в себя биохимические реакции синтеза органических веществ с использованием энергии, накопленной на светозависимой стадии. Чаще всего в качестве таких реакций рассматривается цикл Кальвина и глюконеогенез (образование сахаров и крахмала из углекислого газа воздуха).

Поглощение света пигментами. Электронно-возбужденные состояния пигментов (синглетное, триплетное). Типы дезактивации возбужденных состояний. Наиболее устойчивы те состояния атомов и молекул, в которых валентные электроны занимают самые низкие энергетические уровни и распределены по ним согласно принципу Паули (не более двух электронов с антипараллельными спинами на каждой орбитали). Такое состояние молекулы называют основным синглетным (So) энергетическим состоянием. Суммарный спин всех электронов молекулы в этом случае равен нулю.
Если у возбужденного электрона сохраняется то же направление спина, то молекула находится в возбужденном синглетном состоянии (S*).
Если при переходе на более высокую орбиталь спин электрона обращается, то такое возбуждение называется триплетным (Т*).
Молекула в возбужденном состоянии оказывается способной вступать в реакцию, которая была для нее невозможна, когда она находилась на более низком энергетическом уровне.
Структура молекулы хлорофилла хорошо приспособлена к своим функциям сенсибилизатора фотохимических реакций. В ее состав входят 18 делокализованных π-электронов, что делает молекулу хлорофилла легко возбудимой при поглощении квантов света. Поглощение молекулой хлорофилла света в сине-фиолетовой части спектра обусловлено системой конъюгированных одинарных и двойных связей порфиринового кольца. Поглощение в красной области связано с гидрированием двойной связи у С7-С8 в IV пиррольном ядре (при переходе от протохлорофиллида к хлорофиллиду) и присутствием магния в порфириновом кольце.
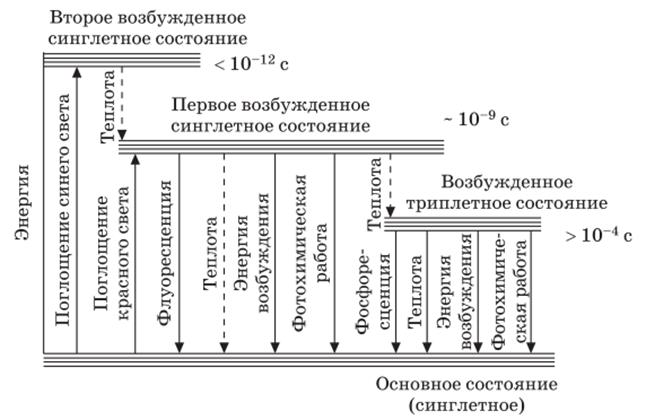
Переходы между возбужденными состояниями хлорофилла
после поглощения кванта красного и синего света
Поглощение молекулой хлорофилла кванта красного света приводит к синглетному электрон-возбужденному состоянию – S1*.
При поглощении кванта синего света с более высоким уровнем энергии электрон переходит на более высокую орбиту (S2*).
Возбужденная молекула хлорофилла возвращается в основное состояние различными путями:
− отдав часть энергии в виде теплоты (особенно с уровня S1*), молекула может излучить квант света с большей длиной волны (правило Стокса), что проявляется в виде флуоресценции. Время жизни синглетного возбужденного состояния 10−3 -10−9 с;
− молекула может из синглетного возбужденного состояния перейти в метастабильное триплетное (с обращением спина) (энергия теряется в виде теплоты). Время жизни триплетного возбужденного состояния >10−4 с;
− из триплетного состояния молекула может вернуться в основное, излучив еще более длинноволновый (чем в случае флуоресценции) квант света. Это обычно более слабое свечение и есть фосфоресценция;
− энергия возбужденного состояния может быть использована на фотохимические реакции. В этом случае флуоресценции и фосфоресценции хлорофилла не наблюдается;
− из первого синглетного и триплетного состояний молекула может переходить в основное с переносом энергии на другую молекулу.
Фотосистемы. Фотосинтетическая единица. Миграция энергии в системе фотосинтетических пигментов. Фотосинтетические пигменты в мембранах хлоропластов имеют не беспорядочное расположение, а организованы в две пигментные системы – фотосистему I (ФС I) и фотосистему II (ФС II).

Фотосистема – совокупность светособирающего комплекса (ССК), фотохимического реакционного центра (РЦ) и переносчиков электрона (цитохромный комплекс).
Хлорофилл выполняет функции: поглощения и передачи энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих (антенных) комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II.
Помимо хлорофилла в ССК имеются каротиноиды, роль которых заключается в поглощении света тех длин волн, которые хлорофилл поглощает сравнительно слабо. ССК растений расположен в мембранах тилакоидов. В каждом ССК содержится от 120 до 240 молекул хлорофиллов, отношение хлорофилла а к хлорофиллу b составляет 1,2-1,4.
Антенный белковый комплекс ФС I содержит 50 молекул хлорофиллов а с максимумами поглощения 680 – 695 нм на один П700 реакционного центра и β-каротин.
Антенный белковый комплекс ФС II содержит 40 молекул хлорофиллов а с максимумами поглощения 670 – 683 нм на один П680 реакционного центра (пигмент с максимумом поглощения 680) и β-каротин.

Пигменты антенных комплексов не обладают фотохимической и энзиматической активностью. Продолжительность синглетного возбужденного состояния исчисляется 10-12 – 10-9 с, и даже на прямом солнечном свету 1 квант света поглощается молекулой хлорофилла не чаще одного раза за 0,1 с. Большую часть времени молекула хлорофилла «простаивает». Поэтому роль пигментов антенных комплексов состоит в том, чтобы собирать и передавать энергию квантов на небольшое количество молекул (молекулы-ловушки) реакционных центров П680 и П700 которые и осуществляют фотохимические реакции. Аналогично отдельные капли дождя ударяют в крышу и, сливаясь, создают постоянный ток воды в водостоке.
Передача энергии идет резонансным путем (механизм Ферстера) и занимает для одной пары молекул 10-10-10-12 сек., расстояние на которое осуществляется перенос составляет около 1 нм.
Природа индуктивного резонанса. Каждая молекула хлорофилла, поглотившая квант света и перешедшая в синглетное возбужденное состояние, является молекулярным осциллятором. Возникающее вокруг возбужденной молекулы переменное электрическое поле с определенной частотой колебаний индуцирует осцилляцию диполя (электрон - ядро) соседней молекулы. При этом молекула-донор переходит в основное состояние, а молекула-акцептор – в возбужденное.
Из первого синглетного и триплетного состояний молекула может переходить в основное
− с выделением энергии в виде света (флуоресценция) или тепла,
− с переносом энергии на другую молекулу пигмента (реализуется в ССК),
− либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносов электрона на другое соединение. Реализуетсяв реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона (восстановителем) и передает его на первичный акцептор.
Чтобы предотвратить возвращение электрона на положительно заряженный хлорофилл, первичный акцептор передает его вторичному. Кроме того, время жизни полученных соединений выше, чем у возбужденной молекулы хлорофилла. Происходит стабилизация энергии и разделение зарядов.
Резонансная миграция энергии между молекулами пигментов разных видов называется гетерогенной (например, хлорофилл – каротиноиды), а перенос энергии между тождественными молекулами хлорофилла а – гомогенной.

Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каротиноидов к хлорофиллу), из-за чего передача возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причем наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
Перенос энергии в антенном комплексе ФСII:
каротин (400–550 нм) → хлорофилл b (650 нм) →
|
|
|
