 |


Зависимость фотосинтеза от условий внешней среды
|
|
|
|
Принцип лимитирующих факторов формулируют следующим образом: при одновременном влиянии нескольких факторов скорость химического процесса «тормозится» фактором, который ближе всех к минимальному уровню (изменение именно этого фактора будет непосредственно влиять на данный процесс).
Этот принцип впервые был установлен Ф. Блекманом в 1915 г. С тех пор было неоднократно показано, что разные факторы, например концентрация СО2 и освещенность, могут взаимодействовать между собой и лимитировать процесс, хотя часто один из них все же главенствует. Освещенность, концентрация СО2 и температура – вот те главные внешние факторы, влияющие на скорость
фотосинтеза. Однако большое значение имеет водный режим, минеральное питание и др.
Одни из указанных факторов, например освещенность и снабжение СО2, действуют на фотосинтез прямо, другие – содержание воды и минеральных элементов в почве - нередко опосредованно, через воздействие на другие физиологические процессы, на темновое дыхание и фотодыхание.
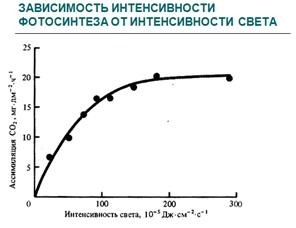
Для большинства растений прямая зависимость интенсивности фотосинтеза от интенсивности света проявляется лишь при сравнительно небольшой величине последнего. В дальнейшем при повышении освещенности прирост фотосинтеза становится все меньше и меньше, и, наконец, кривая фотосинтеза переходит на плато. Это состояние, называемое световым насыщением (СН), свидетельствует о том, что интенсивность фотосинтеза в данный момент больше всего зависит от других, нежели освещенность факторов.
 Все растения по отношению к свету делятся на две большие группы: светолюбивые и теневыносливые. Под светолюбием обычно понимается способность растений расти и развиваться только при хорошем освещении. Светолюбивые травянистые растения не выносят затенения и растут на открытых местах. К ним относятся все сельскохозяйственные культуры, растения лугов, степей, пустынь, солончаков. Среди древесных пород светолюбивыми являются лиственница, сосна, ясень, осина, береза, дуб и др. Они растут на открытых местах или в первом верхнем ярусе леса. Светолюбивые деревья отличаются ажурной кроной, быстрым очищением ствола от сучьев, ранним изреживанием древостоя. Естественное возобновление в таких лесах, как правило, отсутствует или представлено слабо.
Все растения по отношению к свету делятся на две большие группы: светолюбивые и теневыносливые. Под светолюбием обычно понимается способность растений расти и развиваться только при хорошем освещении. Светолюбивые травянистые растения не выносят затенения и растут на открытых местах. К ним относятся все сельскохозяйственные культуры, растения лугов, степей, пустынь, солончаков. Среди древесных пород светолюбивыми являются лиственница, сосна, ясень, осина, береза, дуб и др. Они растут на открытых местах или в первом верхнем ярусе леса. Светолюбивые деревья отличаются ажурной кроной, быстрым очищением ствола от сучьев, ранним изреживанием древостоя. Естественное возобновление в таких лесах, как правило, отсутствует или представлено слабо.
|
|
|
Теневыносливые древесные растения, такие как ель, пихта, клен, вяз, липа, лесная яблоня, рябина и др., хорошо переносят затенение и встречаются как в верхнем ярусе, так и во втором. Высокой теневыносливостью отличаются многие кустарники (лещина, крушина, бересклет, бузина и др.), а также лесные травы (сныть, копытень, ясменник, папоротники, медуница и др.) и мхи. Некоторые их этих трав настолько приспособились жить под пологом леса, что не выносят прямого света и после рубки леса погибают (кислица, недотрога и др.), так как они являются гигрофитами. Эту небольшую группу растений иногда называют тенелюбами. При выставлении таких растений на свет у них резко нарушаются водный режим, фотосинтез и другие жизненно важные процессы.
Теневыносливые древесные растения отличаются от светолюбивых растений густой и плотной кроной, их листья могут развиваться не только по периферии, но и в глубине кроны. Кроме того, крона их имеет обычно большую протяженность по высоте ствола, так как деревья этой группы медленнее очищаются от сучьев. Насаждения из теневыносливых древесных пород характеризуются более медленным самоизреживанием.
Существуют различия анатомической структуры светолюбивых и теневыносливых растений. Листья светолюбивых растений толще, что связано с более сильным развитием палисадной паренхимы (2 - 3 слоя), имеют более плотную сеть жилок. Клетки их листьев обычно меньших размеров, но число устьиц на единицу поверхности значительно больше, благодаря чему СО2 быстрее проникает внутрь листа.
|
|
|
Даже листья одного и того же дерева, располагаясь на различных сторонах кроны, внутри или на периферии ее, имеют явно выраженный световой или теневой характер. Теневые листья отличаются не только структурой, но и большим содержанием хлорофилла, благодаря чему способны использовать небольшое количество света, проникающего в крону.
Для теневыносливых древесных растений характерна листовая мозаика, когда листья отдельных ветвей располагаются в одной плоскости, не затеняя друг друга.
Чем глубже лист расположен в кроне или дальше от вершины, тем меньше он получает света. У светолюбивых древесных растений (лиственница, береза, сосна) побеги желтеют и усыхают при 10 - 20 % от общего количества падающего света, а у теневыносливых при значительно более низких величинах (у ели, пихты, бука - при 1- 3 %, у кипариса - при 0,5 %).
С4-растения относятся к наиболее светолюбивым растениям. Под пологом леса способны расти только теневыносливые С3-растения. Вместе с тем под пологом широколиственных лесов может расти особая группа светолюбивых раноцветущих растений, так называемые эфемероuды, которые способны пройти весь цикл онотогенетического развития до распускания листьев древесных пород, пока освещенность велика (пролеска сибирская, ветреница, гусиный лук, хохлатки).
Минимум освещенности, при которой еще возможно существование древесных растений, выражается следующими величинами: лиственница - 6000 лк, береза - 4000, осина - 3700, сосна - 3300, дуб - 1500, ель - 1000, липа - 700, бук - 400, самшит - 300 лк. Первые в этом ряду древесные растения относятся к наиболее светолюбивым растениям, последние - к наиболее теневыносливым.
Теневыносливые и светолюбивые древесные растения различаются и световым компенсационным пунктом (СКП), под которым понимается та освещенность, при которой растение в процессе фотосинтеза образует столько органических веществ, сколько оно тратит за это же время на дыхание. Растение может расти только в том случае, если освещенность выше СКП. Теневыносливые растения растут при слабой освещенности, прежде всего потому, что обладают невысокой интенсивностью дыхания и соответственно низким СКП. При слабом освещении светолюбивые растения работают с отрицательным балансом, т.е. больше выделяют СО2, чем поглощают, а теневыносливые растения - с положительным.
|
|
|
Приспособление к той или иной освещенности тесно связано с возрастом растения. Всходы древесных растений более теневыносливы, чем взрослые экземпляры.
Световой компенсационный пункт наименьший у теневыносливых и самый высокий у С4- растений. Среднее положение занимают светолюбивые С3-растения. Это свидетельствует о том, что дыхание с наименьшей интенсивностью протекает у теневыносливых растений, затем у светолюбивых С3-растений и, наконец, у растений с С4-типом фотосинтеза. На СКП влияют также условия увлажнения почвы, содержание СО2 в воздухе и некоторые другие факторы. У очень молодых листьев СКП сдвигается в сторону больших значений, ибо у них весьма высока интенсивность дыхания. В целом улучшение факторов среды приводит к сдвигу СКП влево, в сторону меньших значений. Абсолютное значение СКП колеблется от нескольких десятков люкс у теневыносливых, до сотен - у светолюбивых и до 1000-1300 лк у С4-растений.
Связь интенсивности фотосинтеза с освещенностью принято выражать световыми кривыми. Они характеризуются двумя основными показателями: углом наклона кривых к оси абсцисс и точкой СН.
Теневыносливые растения или теневые листья быстрее достигают точки СН, чем другие группы растений и световые листья. Уже при освещенности 5-10 тыс. лк теневыносливые растения достигают максимальных значений фотосинтеза и, несмотря на дальнейшее повышение освещенности, не меняют его. Мало того, при длительном воздействии высокой интенсивности света может наблюдаться снижение показателей фотосинтеза, вызванное фотоокислением хлорофилла и нарушением гранулярной структуры хлоропластов, водного и теплового баланса листьев.
|
|
|
У С4-растений даже при самой высокой освещенности на Земле, равной 70 - 85 тыс. лк, СН не наступает. При более низких значениях освещенности они также имеют наиболее высокую интенсивность фотосинтеза по сравнению с С3-растениями. Очевидно, что деятельность ФЕП-карбоксилазы и при очень сильном освещении «идет в ногу» со световыми реакциями фотосинтеза, и, следовательно, цикл Кальвина работает менее совершенно, чем механизм Хэтча-Слэка у C4-растений.
Своеобразие световых кривых фотосинтеза меняется с возрастом дерева, различиями лесорастительных условий, фактором времени и т.д. Так, например, хвоя ели с молодых деревьев имеет более высокие значения СН, а сами световые кривые располагаются значительно выше таковых для хвои деревьев более старшего возраста. Световые кривые фотосинтеза первичных листьев всходов древесных растений более характерны для листьев светового типа, а семядолей - теневого. Неблагоприятные лесорастительные условия снижают положение СН, свидетельствуя об ослаблении реакции листового аппарата на повышение интенсивности света. Воздействие фактора времени проявляется в том, что длительное пребывание древесных растений (5 -7 лет) в условиях довольно «мягкого» затенения (освещение более высокое, чем освещение СКП) приводит к гибели светолюбивых древесных пород, хотя кратковременное затенение к серьезным нарушениям фотосинтеза и других процессов и структур не ведет.
Угол наклона световых кривых фотосинтеза обычно наиболее высок у теневыносливых растений. Этот показатель используется главным образом для создания математических уравнений связи фотосинтеза с урожайностью и продуктивностью растений.
СКП широко используется при оценке качества и состояния естественного возобновления в лесу, ибо подрост начинает страдать и отмирать под пологом леса при освещенности, равной СКП или ниже его. Он используется также и при назначении времени и интенсивности рубок ухода за лесом в формирующихся молодняках, когда освещенность внутри крон сомкнувшегося полога приближается к СКП. Деревья начинают мешать друг другу взаимным затенением, прирост их падает. Периодичность рубок ухода за лесом также определяется величиной СКП. Этот показатель хорошо объясняет процесс очищения ствола от сучьев. Дело в том, что ветки в кроне дерева начинают отмирать, когда здесь освещенность снижается до СКП или ниже его и когда листья работают с отрицательным балансом, Т.е. больше потребляют ассимилятов от листьев других ветвей, чем их создают сами. При этом качество древесины ствола деревьев улучшается, ибо в густом насаждении формируется бессучковая древесина.
|
|
|
С каждым годом все шире используется способ выращивания растений при искусственном освещении, получивший название светокультуры. Важное значение он имеет в теоретическом и особенно в практическом отношении. Многие теоретические вопросы физиологии и селекции растений решаются с помощью этого способа. Селекционер, используя светокультуру растений закрытого грунта, может за один год получить несколько поколений однолетних культур, что чрезвычайно важно, особенно с экономической точки зрения. В теплицах сейчас получают высокие урожаи овощей, особенно огурцов, томатов, бахчевых культур, а также цветов.
Весьма перспективна и светокультура древесных растений. В опытах В древесные растения давали до 6, а дуб - до 10 приростов за один год. Лиственница при этом усиливала рост в 15 раз. Дуб на непрерывном искусственном свету за один год достигал высоты 1,5 м.
Меняя продолжительность искусственной подсветки, можно получить у сеянцев древесных растений отложение в один год трех годичных слоев древесины. Растения шиповника в условиях светокультуры зацветали на второй год. Но самое важное заключалось в том, что «привычка» давать по несколько приростов в год у саженцев сохраняется и при пересадке их в открытый грунт на естественное освещение. Плодоношение таких древесных растений наступало на несколько лет раньше обычного срока. Перспективна светокультура растений в сочетании с подкормкой углекислым газом. Положительно сказалось на росте и ночное досвечивание сеянцев древесных растений, растущих в открытом грунте. Такая подсветка обычными электрическими лампами накаливания резко повышала прирост сеянцев.
Большое влияние на фотосинтез оказывает спектральный состав света. Еще К.А. Тимирязев установил, что наибольшая интенсивность фотосинтеза наблюдается в красных лучах спектра, затем - в сине-фиолетовых. Минимальный фотосинтез отмечался в зеленых лучах, ибо они почти не поглощаются пластидными пигментами.
Спектральный состав света влияет не только на интенсивность фотосинтеза, но и на качество образующихся в процессе фотосинтеза органических веществ. Так, в частности, красная часть спектра солнечного света в большей степени обусловливает углеводную направленность фотосинтетических продуктов, а сине-фиолетовая часть спектра - белковую.
Очень важным экологическим фактором, влияющим на фотосинтез, является температура воздуха. Необходимо указать, что температура влияет не только на ферментативные реакции этого процесса, но и на темновое дыхание. При повышении температуры до определенных значений, разных для различных видов и даже сортов растений, интенсивность дыхания повышается в большей степени, чем интенсивность фотосинтеза. Подтверждением этого служат экспериментальные данные для некоторых видов американских сосен: отношение интенсивности фотосинтеза к интенсивности дыхания при разных температурных условиях оказалось равным: при температуре 20°С - 13, при 30°С - 7, при 40°С - всего лишь 3.
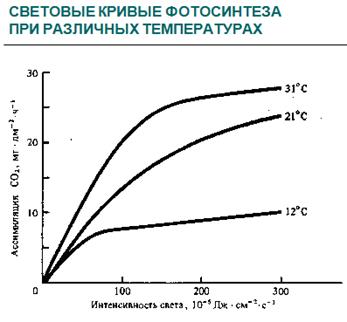 При температуре выше оптимальной (35 - 40 °С) фотосинтез С3-растений снижается из-за нарушения согласованности действия фотосистем, а при 45 – 50 °С он полностью прекращается вследствие инактивации ферментов цикла Кальвина-Бенсона.
При температуре выше оптимальной (35 - 40 °С) фотосинтез С3-растений снижается из-за нарушения согласованности действия фотосистем, а при 45 – 50 °С он полностью прекращается вследствие инактивации ферментов цикла Кальвина-Бенсона.
Вместе с тем для С4-растений такая температура, по существу, является оптимальной, свидетельствуя о приспособлении этих растений к жаркому тропическому климату; коагуляция ферментов наблюдается у них лишь при температуре 60 – 65 °С. Тропические злаки и другие растения с С4-типом фиксации углекислоты отличаются повышенной интенсивностью не только фотосинтеза и газообмена, но и других физиологических процессов: водного режима, минерального питания, обмена органических веществ и т.д. Преобладают у них синтетические процессы над гидролитическими процессами, что в целом предопределяет их очень высокую биологическую и хозяйственную продуктивность.
Фотосинтез при температуре около нуля градусов и ниже ингибируется из-за снижения скорости ферментативных темновых реакций, которые тормозятся в значительно большей степени, чем световые, почти не зависящие от температуры. Естественно, интенсивность фотосинтеза в этих условиях значительно ниже, чем при оптимальной температуре.
В Баварии отмечался положительный фотосинтез у ели в самом конце осени и начале зимы при небольших отрицательных температурах на протяжении большей части дня. Полностью прекращался фотосинтез при температуре воздуха -6 или -8 °С. Для американских сосен и елей также подтверждена возможность зимнего фотосинтеза при -6 °С. При такой же температуре зарегистрирована ассимиляция СО2 у вечнозеленых средиземноморских древесных растений. В Италии с ее теплым климатом эти древесные растения способны к осуществлению фотосинтеза в течение всей зимы. Возможность упомянутого зимнего фотосинтеза связана, очевидно, с тем, что даже в зимний период энергия светового потока поглощается пластидными пигментами древесных растений и концентрируется в хлоропластах с двойной мембраной, согревая их.
У хвойных древесных растений после морозных ночей в ранневесеннее или позднеосеннее время наблюдаемый фотосинтез на следующий теплый день полностью до первоначальной величины не восстанавливается, а дыхание в течение нескольких часов после заморозка резко возрастает, в связи с чем происходит выделение СО2 на свету. Степень снижения интенсивности фотосинтеза находится в прямой зависимости от продолжительности и силы ночного заморозка.
В процессе своей длительной эволюции растения приспособились осуществлять фотосинтез при содержании СО2 в воздухе 0,03 %, или 0,57 мг/л. Вместе с тем на заре возникновения зеленых растений углекислотыбыло значительно больше, что, очевидно, не осталось бесследным для растений. Дополнительные углекислотные подкормки растений в закрытом грунте резко активизируют фотосинтез, а с ним и урожайность. Углекислотное насыщение фотосинтеза в кратковременных опытах наступает у большинства растительных организмов при содержании СО2 0,2 - 0,4 %, что на целый порядок выше, чем в обычных условиях.
 Повышение содержания углекислоты в теплицах вокруг листьев зерновых сельскохозяйственных культур приводит к двойному усилению фотосинтеза, а у томатов, огурцов, табака - к четырехкратному. Неудивительно поэтому повышение урожая тепличных культур на 30 % и даже в 2 раза. Весьма положительно сказываются углекислотные подкормки и при выращивании посадочного материала древесных пород в теплицах. Здесь фотосинтез всходов хвойных древесных растений при подкормке СО2 превышает контроль в 2 раза, а общая биомасса - в 2,7 раза. Вместе с тем растения с С4- типом фотосинтеза на углекислотные подкормки не реагируют. Прямая зависимость фотосинтеза этих растений от содержания СО2 проявляется лишь при концентрациях, существенно более низких, чем естественные. Накапливая большое количество 4-х углеродных органических кислот, С4-растения создают своеобразное «депо» внутренней СО2.
Повышение содержания углекислоты в теплицах вокруг листьев зерновых сельскохозяйственных культур приводит к двойному усилению фотосинтеза, а у томатов, огурцов, табака - к четырехкратному. Неудивительно поэтому повышение урожая тепличных культур на 30 % и даже в 2 раза. Весьма положительно сказываются углекислотные подкормки и при выращивании посадочного материала древесных пород в теплицах. Здесь фотосинтез всходов хвойных древесных растений при подкормке СО2 превышает контроль в 2 раза, а общая биомасса - в 2,7 раза. Вместе с тем растения с С4- типом фотосинтеза на углекислотные подкормки не реагируют. Прямая зависимость фотосинтеза этих растений от содержания СО2 проявляется лишь при концентрациях, существенно более низких, чем естественные. Накапливая большое количество 4-х углеродных органических кислот, С4-растения создают своеобразное «депо» внутренней СО2.
Искусственное увеличение содержания СО2 выше ее насыщающих концентраций уже тормозит фотосинтез С3-растениЙ. Ингибирующее влияние на фотосинтез избытка СО2связывают с повышением кислотности внyrpиклеточных жидкостей, вызывающей смещение их буферности, а также с наркотическим действием углекислоты.
В природе недостаток СО2 проявляется в полдень, когда она усиленно потребляется на фотосинтез. Над фотосинтезирующими листьями содержание СО2 снижается на 12 - 20 % и более. Слабый ветер, приносящий новые порции СО2, усиливает усвоение углерода. Максимальных значений концентрация СО2 под пологом леса достигает к утру. Наибольшее количество углекислого газа отмечается в лиственных и смешанных лесах. В кронах деревьев минимальное содержание СО2 отмечено в летнее время, максимальное - осенью. В нижних слоях воздуха под лесом содержание углекислоты повышается до 0,08 %, что в определенной мере компенсирует недостаток здесь для подроста света.
По аналогии со световыми кривыми фотосинтеза у углекислотных кривых имеются три показателя, в том числе углекислотный компенсационный пункт (УКП) - концентрация СО2, при которой фотосинтез равен дыханию. Для многих древесных растений УКП фотосинтеза чуть больше 0,001 %. У молодых листьев вследствие очень интенсивного дыхания УКП очень высокий и приближается к 0,03 % СО2. Повышенная температура и недостаток воды повышают УКП. С усилением света этот показатель сначала растет, а затем стабилизируется, очевидно, за счет фотодыхания. УКП фотосинтеза С4-растений чрезвычайно низок. Они могут использовать на фотосинтез СО2 до почти нулевой концентрации.
Следует отметить, что в ночное время в растениях накапливается большое количество внутренней СО2. Показано, что растения с С4-типом фиксации СО2 используют этот ее внутренний источник значительно быстрее и эффективнее.
В тканях растений содержится большое количество СО2. Межклетники болотных и водных растений накапливают до 20 %, а древесина древесных растений до 18 % углекислоты. Такие высокие концентрации этого газа не вызывают, очевидно, нарушений физиологических процессов. Внутритканевая углекислота - дополнительный источник углеродного питания растений. На процесс фотосинтеза может использоваться не только СО2 атмосферы, внутренняя СО2 и СО2 4-х углеродных органических кислот после их декарбоксилирования у растений с С4- и САМ типами фотосинтеза, но и СО2 почвы и почвенного раствора.
Содержание СО2 в атмосфере, начиная с конца XVIII века, стало повышаться сначала из-за чрезмерной вырубки лесов, а затем за счет промышленной углекислоты. В 1860 г. атмосфера содержала 0,0295 %, в 1979 - 0,0335 % СО2. По прогнозам, к концу нашего столетия концентрация СО2 в воздухе достигнет 0,038 - 0,040 %, а к 2025 г. - 0,053 - 0,070 %, Т.е. возрастет в среднем в 2 раза. В кратковременных опытах такое повышение содержания СО2 приводит к адекватному усилению фотосинтетической активности и роста растений. Длительные же эксперименты как с травянистыми, так и с древесными растениями в значительной степени противоречат этому. Так, у сеянцев сосны желтой появились признаки угнетения уже через год, а у некоторых даже через полгода пребывания их в атмосфере с 0,06 % СО2: наблюдалось резкое снижение содержания хлорофиллов а и b, каротиноидов, обнаруживалась пятнистость хвои, которая быстро старела и опадала. Пребывание сеянцев кедра атласского и сосны черной в течение года при таком же уровне углекислоты не приводило к повышению интенсивности фотосинтеза, но вызывало увеличение прироста в высоту и по диаметру на 20 и 10 % соответственно. Двойное увеличение концентрации СО2 в воздухе у саженцев тополя евроамериканского уже через 2 недели снизилась интенсивность фотосинтеза в 2 раза, но несколько увеличился размер листовой пластинки.
Таким образом, при длительном нахождении растений в условиях с повышенным содержанием СО2 происходит затоваривание ею клеток мезофилла, уменьшается размер зияния устьиц, снижается активность ключевого фермента - карбоксилазы РДФ, замедляется регенерация РДФ, основного акцептора углекислоты и т.д. Некоторое усиление ростовых процессов может быть связано с феноменом кислого роста, проявляющегося под влиянием повышенных концентраций СО2. В данном случае снова имеем дело с фактором времени. Для ответа на вопрос, может ли растительность, в том числе лесная, ослабить усиление парникового эффекта, нужны дополнительные длительные опыты непосредственно в природной обстановке по фотосинтезу и росту древесных растений в условиях обогащенной СО2 атмосферы и при сочетании их с другими естественными (засуха, засоление, недостаток питательных веществ в почве и др.) и антропогенными стрессами (промышленные загрязнения, рекреация и т. д.).
Известно, что доля СО2 в создании парникового эффекта на Земле составляет 43 %, метана 23 % и закиси азота 3 %. Следовательно, СО2 является главным парниковым газом. Повышение ее содержания может привести к изменению климата на планете. По прогнозам, средняя температура воздуха к 2025 г. возрастет на 1,8 - 2,5 °С, а уровень мирового океана только за счет теплового расширения воды поднимется на 65 см, что вместе с водой тающих вечных снегов и льдов приведет к затоплению и засолению огромной площади прибрежных территорий. Вместе с этим усилится аридность многих регионов мира, произойдут изменения в распределении растительности и составе фитоценозов. Повышенная температура почвы может отрицательно сказаться на процессах покоя семян древесных растений, их прорастании и формировании всходов и подроста, т.е. отрицательно скажется на процесс ах естественного возобновления леса.
Человечество обеспокоено наступлением экологического кризиса на Земле. Научная общественность требует совершенствования промышленных технологий по принципу замкнутых циклов, позволяющих достаточно полно утилизировать промышленные выбросы, в том числе СО2 и т.д. Лесоводами разрабатываются программы по созданию рукотворных лесов из быстрорастущих древесных пород с целью большего вовлечения промышленной СО2 в фотосинтетический цикл.
Специфически действует на интенсивность фотосинтеза ветер. Равномерный, со скоростью примерно 0,5 - 1,1 м/с ветер способствует притоку СО2 из других мест к фотосинтетической поверхности полога леса и тем самым повышает фотосинтез. Сильный же ветер, увеличивая водный дефицит листа, действует на фотосинтез уже угнетающе. Вместе с тем растения достаточно приспособлены к действию ветра. Хорошо выраженная подвижность черешка листьев осины и некоторых других древесных пород, связанная с особенностями его строения, дает возможность листьям соприкасаться с большим объемом воздуха и СО2, что помогает этим древесным породам даже при очень слабом движении воздуха осуществлять фотосинтетический процесс с достаточно высокой интенсивностью.
Процесс фотосинтеза очень чувствителен к изменению водного режима почвы и зависит от доступности почвенной воды корням растений. Наиболее благоприятной для абсолютного большинства растений является влажность почвы, составляющая 60-70 % от полной влагоемкости. Содержание воды выше и ниже указанного уровня снижает интенсивность фотосинтеза. Обезвоживание клеток листа приводит к закрыванию устьиц и увеличению сопротивления диффузии СО2 внутрь листа, а также к инактивированию ферментов (синтетаз).
Содержание воды в листе оказывает большое влияние на фотосинтез. При небольшом водном дефиците (до 5 %) интенсивность фотосинтеза может достигать максимальных значений. Более сильное обезвоживание тканей листа вызывает снижение интенсивности фотосинтеза, а затем падение ее до нуля. Полное насыщение клеток мезофилла листа водой также несколько снижает скорость этого процесса.
В природных условиях недостаток воды в почве наиболее часто снижает интенсивность фотосинтеза. Особенно вредно отражается на фотосинтезе комплексное воздействие почвенной и атмосферной засухи. Различные древесные породы реагируют на это по-разному. Так, дуб черешчатый в меньшей степени снижает интенсивность фотосинтеза в период засухи, чем клен остролистный и ясень пушистый.
По ходу вегетации в большинстве районов с умеренным климатом древесные растения испытывают недостаток воды в почве на протяжении большего или меньшего периода времени. Поэтому после дождя или полива интенсивность фотосинтеза деревьев повышается. После непродолжительной засухи первоначальная активность фотосинтеза обычно восстанавливается в течение нескольких дней. Недостаток воды в почве, снижая фотосинтез прямо, а также косвенно через уменьшение площади листьев, влияет отрицательно на общую фотосинтетическую продуктивность растений.
Сущность вредного влияния на фотосинтез избытка воды в почве сводится к тому, что вода вытесняет из почвы воздух, а вместе с ним и кислород. В затопленной почве создаются полуанаэробные или анаэробные условия. Корни растений не могут в связи с этим поглощать воду в достаточных количествах, и в листьях возникает водный дефицит. Снижение интенсивности фотосинтеза под влиянием затопления почвы, особенно застойной водой, наблюдалось как у плодовых, так и у лесных лиственных и хвойных древесных растений. Постоянный избыток воды в почве в большей степени ингибирует этот процесс, чем периодический. На торфяных почвах таежной зоны убедительно показано положительное влияние осушения на фотосинтез древесных растений.
Достаточно чувствителен фотосинтез и к обеспечению растений элементами минерального питания. К настоящему времени доказано многообразное действие недостатка или избытка макро- и микроэлементов на активизацию или блокирование деятельности ферментных систем растений, ответственных за отдельные реакции фотосинтеза, их скорость и направленность, биосинтез зеленых пигментов и каротиноидов и формирование хлоропластов. Весьма сильно косвенное влияние минеральных элементов на фотосинтез через воздействие на процессы транспорта ассимилятов и отложение веществ в запас, дыхание, водный режим и состояние цитоплазмы клеток мезофилла листа, движение устьиц, размеры листовой пластинки и структуру фотосинтетического аппарата. Сильное отрицательное влияние на фотосинтез оказывает недостаток в листьях, прежде всего азота, затем фосфора и калия.
Так, снижение в листьях тополя содержания азота с 3,8 до 1,8 % уменьшало фотосинтез на одну треть; при этом световое насыщение у бедных азотом листьев наступало при 20 тыс. ЛК, тогда как у богатых азотом листьев этого не наблюдалось даже при 40 тыс. лк. Если учесть, что при недостатке азота поверхность листа составляла всего лишь 45 % таковой у деревьев тополя, богатых азотом, то различия в общей фотосинтетической продуктивности сравниваемых объектов окажутся еще более внушительными.
Значительное число исследований выполнено по фотосинтезу плодовых древесных пород на различном фоне минерального питания. Например, фотосинтез листьев яблони с ярко выраженным недостатком азота был почти в 3 раза меньше по сравнению с удобренным контролем. Показано также, что интенсивность фотосинтеза снижалась не только из-за недостатка минеральных элементов, но и от избытка одного из них.
Минеральные элементы, находясь в минимуме, не только снижают интенсивность процесса фотосинтеза, но и отрицательно действуют на прирост фотосинтетической поверхности растения. Эти две причины и ограничивают урожай, и продуктивность растений в условиях с недостатком или дисбалансом минеральных элементов в почве. От гармонической взаимосвязи воздушного (фотосинтез) и корневого питания в значительной мере зависит физиологическое состояние растений.
При выращивании посадочного материала в лесных питомниках и теплицах снабжение молодых растений с первых дней их существования минеральными элементами ведет к более быстрому развитию за счет резкого усиления фотосинтетической активности.
В течение дня у растений интенсивность фотосинтеза изменяется. Утром фотосинтез идет, как правило, с достаточно высокой скоростью из-за очень малого водного дефицита листьев. В умеренную погоду при достаточной влажности почвы и воздуха фотосинтез возрастает постепенно, достигая максимальных значений в полдень. Дневной ход фотосинтеза описывается одновершинной кривой, следуя за изменениями освещенности и температуры. Переменная погода ведет к полной зависимости этого хода от освещенности с образованием многовершинной кривой.
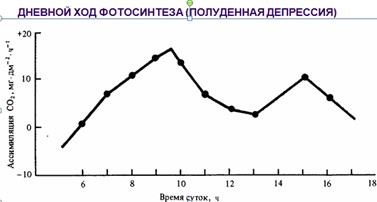 В большинстве же случаев по мере повышения освещенности и температуры интенсивность фотосинтеза достигает максимальных значений перед полуднем, затем наблюдается ее спад с последующим вторым максимумом, образуя двухвершинную кривую. При очень жаркой и сухой погоде кривая становится одновершинной с максимумом в ранние утренние часы. Наблюдающийся спад фотосинтеза получил название полуденной депрессии.. Среди условий, вызывающих эту депрессию, следует назвать водный дефицит листьев, закрывание устьиц, перегрев листьев, слабый отток ассимилятов из хлоропластов, фотоокисление хлорофилла и инактивацию ферментов, в том числе карбоксилаз, на сильном свету, резкое повышение дыхания, понижение содержания СО2 около листьев в связи с ее усиленным потреблением зелеными листьями.
В большинстве же случаев по мере повышения освещенности и температуры интенсивность фотосинтеза достигает максимальных значений перед полуднем, затем наблюдается ее спад с последующим вторым максимумом, образуя двухвершинную кривую. При очень жаркой и сухой погоде кривая становится одновершинной с максимумом в ранние утренние часы. Наблюдающийся спад фотосинтеза получил название полуденной депрессии.. Среди условий, вызывающих эту депрессию, следует назвать водный дефицит листьев, закрывание устьиц, перегрев листьев, слабый отток ассимилятов из хлоропластов, фотоокисление хлорофилла и инактивацию ферментов, в том числе карбоксилаз, на сильном свету, резкое повышение дыхания, понижение содержания СО2 около листьев в связи с ее усиленным потреблением зелеными листьями.
За Полярным кругом в июне и июле в период белых ночей фотосинтез наблюдается круглые сутки. Продолжительность фотосинтеза сеянцев сосны в лесных питомниках под Архангельском в июле составила 18 часов, ели - 18 - 20, что примерно на 3 - 4 часа превышает продолжительность непрерывного фотосинтеза хвойных древесных растений в Центральной Европе.
Эти и другие материалы свидетельствуют о том, что в северных районах малая продолжительность вегетационного периода в значительной мере компенсируется круглосуточным фотосинтезом растений. Здесь, как правило, полуденная депрессия фотосинтеза наблюдается лишь в очень ясные дни с температурой выше 20 °С.
Сезонный ход фотосинтеза лиственных и хвойных древесных растений, за исключением листопадной лиственницы, различен. Хвойные породы с уже имеющимся фотосинтетическим аппаратом раньше начинают активный фотосинтез и позже его заканчивают. Наряду с этим обычно более высокая фотосинтетическая активность лиственных пород в разгар вегетационного периода ведет к тому, что их продуктивность в сравнительно сходных лесорастительных условиях, как правило, не ниже, чем у хвойных пород.
Наряду с естественными факторами на фотосинтез растений оказывают влияние и различные антропогенные воздействия, промышленные газы, пыль, сажа и др.
Фотосинтетический аппарат растений, в том числе древесных, весьма чувствителен к вредным газам. Особенно часто встречаются повреждения, связанные с действием сернистого газа (SО2), который ингибирует транспорт электронов в хлоропластах и фотолиз воды, нарушает цикл Кальвина - Бенсона, замедляет транспорт ассимилятов.
Больше всего газа проникает в период активного фотосинтеза, т.е. когда устьица открыты полностью. Концентрация сернистого газа, равная 0,00004 %, оказалась токсичной для многих видов хвойных растений. Наряду со снижением интенсивности фотосинтеза сернистый газ изменял и качественный состав продуктов фотосинтеза.
Отрицательное влияние на фотосинтез оказывают и другие промышленные газы - окись углерода, хлор, фтор и др. Ингибируют фотосинтез и окислы азота. Вместе с тем депрессия фотосинтеза в отдельных случаях бывает неглубокой и обратимой.
В меньшей степени изучено воздействие на фотосинтез различных пылеватых частиц и тяжелых металлов. Пыль, сажа, металлические частички в воздухе вокруг крупных промышленных предприятий плотным слоем покрывают листья растений, в том числе древесных, почти полностью закупоривают устьица, резко снижают (до двух раз) доступ света к мезофиллу листа. Цементная пыль образует на поверхности листа твердую корку, которая часто механически повреждает покровы листа. Иногда высокие концентрации солей, содержащихся в пыли ряда заводов, приводят к воздушному засолению растений. Вот почему на листьях нередко появляются отмершие участки ткани (некротические области) и тем более обширные, чем менее устойчиво дерево к загрязнениям. Все это резко снижает интенсивность фотосинтеза и повышает дыхательную активность, что отрицательно сказывается на общей продуктивности лесов, подверженных действию твердых пылевых токсикантов.
Значительное ослабление интенсивности фотосинтеза связано также с различными заболеваниями растений, в том числе грибными (ржавчина, мучнистая роса, мильдью и др.).
Так, например фотосинтез листьев дуба черешчатого, пораженных мучнистой росой, протекал на более низком уровне в течение всего дня. Ингибирование фотосинтеза зависит от степени поражения листа. В опытах с трехлетними саженцами той же породы при слабой степени поражения (мицелием занято 10 % поверхности листа), фотосинтез уменьшался на 37 %, а при сильной (поражено более 50 %) - на 71 % по сравнению с непораженными экземплярами.
Ингибирование процесса фотосинтеза под влиянием патогена может происходить вследствие действия выделяемых грибом токсинов, вызывающих деструктивные изменения фотосистем с последующим нарушением ультраструктуры хлоропластов и отмиранием отдельных участков листа, снижения содержания хлорофилла и каротиноидов, затемнения листа мицелием гриба и т.д.
|
|
|
