 |


Окисление воды и образование кислорода происходит в ФС II.
|
|
|
|
При возбуждении Р680 электрон передается на феофитин, а оттуда – на пластохиноны. Дефицит электрона в Хл-а+*, образовавшийся в процессе электронного транспорта, компенсируется за счет электронов, полученных при окислении волы. Катионы марганца и остатки тирозина участвуют в транспорте электронов от воды к хлорофиллу. Радикал Хл-а+* является настолько сильным окислителем, что он способен отнять электрон у остатка тирозина в составе белка реакционного центра; при этом образуется радикал тирозина. Обычно тирозин, участвующий в этой реакции, обозначают как Z. Дефицит электрона в радикале тирозина компенсируется за счет окисления ионов марганца. Комплекс ФС II содержит четыре иона марганца, расположенных близко друг к другу. Эта группа ионов марганца получила название марганцевого кластера. Чтобы образовать в процессе окисления воды одну молекулу O2, реакционный центр должен передать четыре электрона. Марганцевый кластер представляет собой редокс-систему, которая может отдавать последовательно четыре электрона акцептору и затем отбирать их у донора. При этом ионы Мn, очевидно, могут находиться в состояниях Мn3+ и Мn4+.
Образно говоря, четыре электрона, необходимые реакционному центру, берутся «взаймы» у Mn-кластера и затем разом возвращаются к ионам марганца при окислении воды до кислорода. Протоны, высвобождающиеся при фотоокислении воды, поступают в люмен тилакоидов.
2 Ф ОТОСИНТЕТИЧЕСКОЕ ФОСФОРИЛИРОВАНИЕ
Фотохимические процессы в реакционных центрах ФСI и ФСII приводят к быстрому первичному запасанию энергии квантов света в форме лабильных соединений с высоким энергетическим потенциалом. Дальнейшие реакции фотосинтеза направлены на преобразование энергии света в более стабильную форму: сначала в форму восстановленного НАДФН и АТФ, а затем, при использовании этих продуктов в реакциях углеродных циклов, – в форму углеводов и других стабильных органических соединений.
|
|
|
Световые реакции фотосинтеза сопровождаются реакциями образования АТФ. Любой механизм образования АТФ носит название фосфорилирования. В случае фотосинтеза говорят о фотофосфорилировании, подчеркивая, что АТФ образуется за счет трансформации энергии света.
Образование восстановленного НАДФН и АТФ в хлоропластах связано с работой электрон-транспортной цепи (ЭТЦ) фотосинтеза. В зависимости от характера транспорта электронов различают циклическое и нециклическое фотофосфорилирование. Сначала энергия квантов света преобразуется в энергию мембранного электрохимического потенциала, образованного за счет неравномерного распределения протонов на внешней и внутренней сторонах фотосинтетической мембраны, а затем в ходе работы АТФ-синтетазы превращается в химическую – АТФ.
Для восстановления одной молекулы NADP+ в процессе фотосинтеза необходимы два электрона и два протона, причем донором электронов является вода. Фотоиндуцированное окисление воды осуществляет ФС II, восстановление NADP+ – фотосистема I. Следовательно, эти две фотосистемы должны функционировать во взаимодействии.
Исходя из данных о квантовом выходе фотосинтеза (необходимы 8 квантов света для выделения одной молекулы О2) из состава компонентов, входящих в ФС I и ФС II, и величин их окислительно-восстановительного потенциала была разработана схема последовательности реакций в световой фазе фотосинтеза. Из-за сходства с буквой Z эта схема получила название Z-схемы.
В ФС II димер П680, поглотив энергию, эквивалентную 2 квантам коротковолнового красного света, и перейдя в синглетное возбужденное состояние, отдает 2 электрона феофетину (Фф). От Фф электроны, теряя энергию, последовательно передаются на пластохиноны ФС II – QA и QB,, далее на пул липидорастворимых молекул пластохинона (PQ), переносящих через липидную фазу мембраны электроны и протоны, на железосерный белок FeSR и цитохром f цитохромного комплекса b6– f, восстанавливая Cu-содержащий белок пластоцианин (Пц).
|
|
|
Вакантные места («дырки») в П680 заполняются двумя электронами из содержащего Мn-кластер переносчика электронов Z, который в свою очередь восстанавливается за счет фотоокісленія воды.

Однако описанная цепь реакций резко замедляется, если не возбуждена ФС I, так как в этом случае весь Пц переходит в восстановленное состояние. При возбуждении П700 в реакционном центре ФС I энергией, эквивалентной 2 квантам длинноволнового красного света, 2 электрона захватываются мономерной формой хлорофилла a (A1) и затем последовательно передаются переносчикам электронов А2 и Ав (железосерные белки FeS), ферредоксину (водорастворимый FeS-белок) и ферредоксин:NADP-оксидоредуктазе с FAD в качестве кофактора. Наконец, редуктаза восстанавливает NADP +.
На вакантные места в П+700 переходят электроны с Пц, и нециклическая цепь переноса электронов таким образом замыкается.
Энергия, освобождающаяся при движении электронов от П680 до П700, используется для синтеза АТФ из AДФ и неорганического фосфата (фотофосфорилирование).
Таким образом, нециклический транспорт электронов сопровождается выделением кислорода и восстановлением НАДФ+ и сопряжен с синтезом АТФ (нециклическое фотофосфорилирование).
Циклический транспорт электронов осуществляется отдельно в ФС I или ФСII.
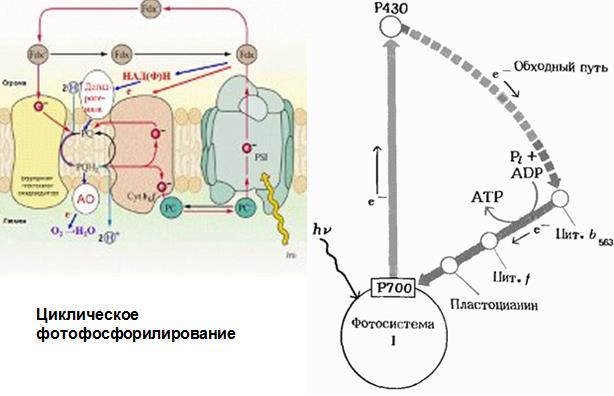
Циклический транспорт в ФС I включает перенос электронов от восстановленого ферредоксина(Фдвост.) обратно к окисленному П+700. В этом случае возбужденные молекулы П700 последовательно отдают электроны на А1, А2, АВ, Фд, PQ, цитохром b6, FeSR, цитохром f, Пц и, наконец, П700. При этом NADP+ не восстанавливается. Освобождающаяся энергия используется для фосфорилирования ADP.
Циклический поток электронов с участием ФСI сопряжен с с синтезом АТФ (циклическое фотофосфорилирование) и может обеспечивать дополнительный синтез АТФ, необходимый для ассимиляции углерода. Он может быть альтернативным путем использования энергии света при его высоких интенсивностях.
|
|
|
Циклический транспорт электронов в ФС II связан с обратным переносом электронов от восстановленных первичных хинонов QA и QB к окисленному пигменту П+680. При этом участвуют цитохром b559, β -каротин и сопровождающие молекулы хлорофилла а РЦ ФСII.

Является альтернативным путем использования энергии света. Он активируется в условиях, когда интенсивность света превышает возможности ЭТЦ утилизировать его энергию или при повреждении водоокисляющей системы хлоропластов.
Итак, основные этапы преобразования энергии, приводящие к образованию АТФ и НАДФ•Н, включают в себя следующие процессы:
1) поглощение энергии света пигментами светособирающей антенны;
2) перенос энергии возбуждения к фотореакционному центру;
3) окисление фотореакционного центра и стабилизация разделенных зарядов;
4) перенос электрона по цепи электронного транспорта, образование НАДФ•Н;
5) трансмембранный перенос ионов водорода;
6) синтез АТФ.
3 Пути фиксации со2 при фотосинтезе
В результате фотохимических реакции в хлоропластах создается необходимый уровень АТР и NADPH. Эти конечные продукты световой фазы фотосинтеза стоят на входе в темновую фазу, где СО2 восстанавливается до углевода:
Сами по себе АТР и NADPH не в состоянии восстановить СО2. Очевидно, и темновая фаза фотосинтеза - сложный процесс, включающий большое количество реакций. Кроме того, существуют различные пути восстановления СО2. В настоящее время известны так называемые С3-путь и С4-путь фиксации СО2, фотосинтез по типу толстянковых (САМ-метаболизм) и фотодыхание.
С3-путь. Этот способ ассимиляции СО2, присущий всем растениям, в 1946-1956 гг. был расшифрован американским биохимиком М. Кальвином и его сотрудниками, в силу чего он получил название цикла Кальвина. Этот цикл, весьма напоминающий обращенный пентозофосфатный путь дыхания, состоит из трех этапов: карбоксилирования, восстановления и регенерации.
1. Карбоксилирование. Молекулы рибулозо-5-фосфата фосфорилируются с участием АТР и фосфорибулозокиназы, в результате чего образуются молекулы рибулозо-1,5-дифосфата, к которым в свою очередь присоединяется СО2 с помощью рибулозодифосфаткарбоксилазы. Полученный продукт расщепляется на две триозы: 2 молекулы 3-фосфоглицериновой кислоты (3-ФГК).
|
|
|
2. Фаза восстановления. 3-ФГК восстанавливается до 3-фосфоглицеринового альдегида (3-ФГА) в два этапа. Сначала происходит фосфорилирование 3-ФГК при участии АТР и фосфоглицераткиназы до 1,3-дифосфоглицериновой кислоты, а затем восстановление 1,3-ФГК с помощью NADPH и дегидрогеназы фосфоглицеринового альдегида.
3. Фаза регенерации первичного акцептора диоксида углерода и синтеза конечного продукта фотосинтеза. В результате описанных выше реакций при фиксации трех молекул СО2 и образовании шести молекул восстановленных 3-фосфотриоз пять из них используются затем для регенерации рибулозо- 5-фосфата, а один - для синтеза глюкозы. 3-ФГА под действием триозофосфатизомеразы изомеризуется в фосфодиоксиацетон. При участии альдолазы 3-ФГА и фосфодиоксиацетон конденсируются с образованием фруктозе-1,6-дифосфата, у которого отщепляется один фосфат с помощью фруктозе-1,6- дифосфатазы. В дальнейших реакциях, связанных с регенерацией первичного акцептора СО2, последовательно принимают участие транскетолаза и альдолаза. Транскетолаза катализирует перенос содержащего два углерода гликолевого альдегида от кетозы на альдозу.
Альдолаза затем осуществляет перенос трехуглеродного остатка фосфодиоксиацетона на альдозу, в данном случае на эритрозо-4-фосфат, в результате чего синтезируется седогептулозо-1,7-дифосфат. Последний дефосфорилируется и под действием транскетолазы из него и 3-ФГА образуются ксилулозо-5-фосфат и рибозо-5-фосфат. Две молекулы ксилулозо-5-фосфата при участии рибулозофосфатэпимеразы и одна молекула рибозо-5-фосфата с участием рибозофосфатизомеразы превращаются в три молекулы рибулозо-5-фосфата, с которого начинается новый цикл фиксации СО2.

Из оставшейся неиспользованной 6-й молекулы 3-ФГА под действием альдолазы синтезируется (при повторении цикла) молекула фруктозе-1,6-дифосфата, из которой могут образовываться глюкоза, сахароза или крахмал:
Таким образом, для синтеза одной молекулы глюкозы в цикле Кальвина необходимы 12 NADPH и 18 АТР, которые поставляются в результате фотохимических реакций фотосинтеза.
С4-путь. С3-цикл – основной, но не единственный путь восстановления двуокиси углерода. Так, австралийские ученые М.Д. Хетч и К.Р. Слэк в 1967 г. обнаружили, что у кукурузы, сахарного тростника, сорго и некоторых других тропических растений первичным продуктом темновой фазы является не ФГК, а ЩУК. Кроме того, было обнаружено, что эти растения имеют не только гранальные, но и агранальные хлоропласта. Первые расположены в основных клетках мезофилла, а вторые – в паренхимных клетках, окружающих одним слоем каждый проводящий пучок. Этот слой клеток был назван обкладкой проводящего пучка.
|
|
|
В результате дальнейших исследований был открыт еще один цикл фотосинтеза, названный циклом Хетча – Слэка, или С4-циклом. Последнее название связано с тем, что ЩУК содержит в своей молекуле 4 атома углерода.
Акцептором углекислого газа в этом цикле является фосфоенолпируват (ФЕП). В результате карбоксилирования ФЕП образуется ЩУК и ортофосфат.
К С4-растениям относится ряд культурных растений преимущественно тропического и субтропического происхождения - кукуруза, просо, сорго, сахарный тростник и многие злостные сорняки - свинорой, сыть округлая, ежовник крестьянский, просо куриное, просо крупное, гумай (сорго алепское), щирица, щетинник и др. Как правило, это высокопродуктивные растения, устойчиво осуществляющие фотосинтез при значительных повышениях температуры и в засушливых условиях.
С4-цикл можно разделить на две стадии: карбоксилирование, происходящее в клетках мезофилла, и декарбоксилирование и синтез углеводов, идущие в клетках обкладки проводящих пучков. Общим для всех С4-растений является то, что карбоксилированию подвергается ФЕП при участии ФЕП-карбоксилазы и образуется ЩУК, которая восстанавливается до яблочной кислоты или аминируется с образованием аспарагиновой кислоты.
Щавелевоуксусная, яблочная и аспарагиновая кислоты являются четырехуглеродными соединениями.
По способу декарбоксилирования при участии НАДФ-Н или НАД-малатдегидрогеназы (МДГ) или ФЕП-карбоксикиназы (ФЕП-КК) у С4-растений можно выделить три группы: НАДФ-МДГ, НАД-МДГ и ФЕП-КК-типы соответственно.
У НАДФ-МДГ-растений главными метаболитами, вовлеченными в обмен между клетками, являются малат и пируват (ПВК), у НАД-МДГ-растений - аспартат и аланин и у ФЕП-КК-растений - аспартат и ФЕП. Важнейшие сельскохозяйственные культуры - кукуруза, сорго, сахарный тростник и такие распространенные сорняки, как сыть, ежовник, щетинник, гумай, относятся к НАДФ-МДГ-типу.
Рассмотрим С4-цикл восстановления СО2 на примере этих растений. СО2, поступающий в лист через устьица, попадает в цитоплазму клеток мезофилла, где при участии ФЕП-карбоксилазы вступает в реакцию с ФЕП, образуя ЩУК. Затем уже в хлоропластах ЩУК восстанавливается до яблочной кислоты (малата) за счет НАДФ-Н, образующегося в ходе световой фазы фотосинтеза; ЩУК в присутствии NH4 может превращаться также в аспартат.

Затем малат переносится в хлоропласты клеток обкладки сосудистого пучка, где он подвергается окислительному декарбоксилированию, продуктом которого является пировиноградная кислота (ПВК). Последняя снова диффундирует в мезофилл, где при участии АТФ, образованной в световой фазе, происходит регенерация ФЕП, после чего цикл карбоксилирования повторяется с участием новой молекулы СО2.
Образовавшиеся в результате окислительного декарбоксилирования малата СО2 и НАДФ-Н поступают в цикл Кальвина, что приводит к образованию ФГК и других продуктов, свойственных С3-растениям. Следовательно, именно клетки обкладки выполняют роль основной ассимилирующей ткани, поставляющей сахара в проводящую систему. Клетки мезофилла выполняют вспомогательную функцию – подкачку СО2 для цикла Кальвина.
Таким образом, С4-путь обеспечивает более полное усвоение СО2, что особенно важно для тропических растений, где основным лимитирующим фактором фотосинтеза является концентрация СО2. Эффективность усвоения СО2 С4-растениями увеличивается также за счет подачи НАДФ-Н в хлоропласты клеток обкладки. Эти хлоропласты имеют агранальное строение и специализируются на темновой фазе фотосинтеза, здесь практически не происходит нециклическое фотофосфорилирование. На один агранальный хлоропласт в среднем приходится 8-10 гранальных хлоропластов, осуществляющих первичную фиксацию СО2 и нециклическое фотофосфорилирование. Такая компартментация процессов и кооперация функционирования тканей обеспечивают повышение продуктивности растений и позволяют накапливать СО2 в органических кислотах для осуществления фотосинтеза даже при закрытых устьицах в наиболее жаркое время дня. Это сокращает потери воды на транспирацию. Эффективность использования воды С4-растениями вдвое выше, чем у С3-растений.
САМ-фотосинтез. В сухих местах существуют растения-суккуленты, у которых устьица открыты ночью и закрыты днем для уменьшения транспирации. Поэтому эти растения поглощают углекислый газ ночью. Поглощаемый СО2 используется, как и у С4-растений, для карбоксилирования ФЕП. ФЕП образуется ночью из крахмала в результате его гидролиза и включения образовавшегося глюкозо-6-фосфата в гликолиз. Катализирует реакцию карбоксилирования ФЕП-карбоксилаза. В результате образуется ЩУК и ортофосфат.

Образовавшаяся ЩУК восстанавливается под действием NADH-зависимой малатдегидрогеназы до яблочной кислоты, которая накапливается в вакуолях клеток листа. Это приводит к закислению клеточного сока в ночное время. Днем в условиях высокой температуры, когда устьица закрыты, малат транспортируется из вакуолей в цитоплазму и там декарбоксилируется при участии декарбоксилирующей малатдегидрогеназы с образованием СО2 и ПВК. СО2 поступает в хлоропласты и включается в них в цикл Кальвина, участвуя в синтезе сахаров.
Таким образом, у растений с фотосинтезом по типу толстянковых много общего с С4-путем фотосинтеза. Однако при САМ-метаболизме фиксация СО2 с образованием малата (ночью) и декарбоксилирование малата с высвобождением СО2 и пирувата (днем) разделены во времени. У С4-растений эти же реакции разграничены в пространстве: первая протекает в хлоропластах мезофилла, вторая — в клетках обкладки.
Фотодыхание
Если освещать растение ярким светом, а затем быстро поместить в темноту, то в первые минуты наблюдается усиление поглощения листьями кислорода и выделение углекислоты. Это светоактивируемое выделение СО2 и поглощение О2 листьями получило название фотодыхания. Оно характерно для растений С3- типа фиксации углекислоты и внешне не обнаруживается у растений с С4-типом.

Если учитывать фотодыхание, то на свету в листьях С3-растений одновременно идут три различных процесса: фотосинтез; митохондриальное (темновое) дыхание; фотодыхание. В связи с этим для получения истинных значений фотосинтеза необходимо принимать во внимание не только темновое дыхание, но и фотодыхание.

Фотодыхание обнаружено и у древесных растений. Этот процесс наблюдался у разных видов сосны, ели, лиственницы, пихты, различных видов и клонов тополя, тюльпанного дерева и других представителей древесного мира.
Фотодыхание осуществляется в особых микротельцах - пероксисомах, имеющих плотную гранулярную строму, окруженную простой мембраной. В них сосредоточены гликолатоксидаза, каталаза, глютамат- и глиоксилаттрансаминаза и другие ферменты, катализирующие превращения веществ в процессе фотодыхания. В ходе этого процесса развертывается довольно сложная картина различных превращений гликолевой кислоты, одного из промежуточных продуктов фотосинтеза.
В присутствии кислорода на свету РДФ-карбоксилаза присоединяет к РДФ не СО2, а О2. Присоединив кислород, РДФ распадается на одну молекулу ФГК и одну молекулу фосфогликолевой кис лоты, которая, отщепляя остаток фосфорной кислоты, превращается в гликолевую и из хлоропластов переходит в пероксисомы. Здесь гликолевая кислота окисляется в глиоксилевую с образованием пероксида. Образующаяся перекись водорода распадается на воду и свободный кислород. Глиоксилевая кислота, реагируя с глутаминовой кислотой, трансаминируется в глицин, который покидает пероксисомы и в митохондриях превращается в другую аминокислоту - серин: из двух молекул глицина образуется одна молекула серина и выделяется одна молекула СО2. Вследствие этого часть зафиксированного в фотосинтезе углерода теряется растением.
Фотодыхание теснейшим образом связано не только с отдельными звеньями фотосинтеза, но и с темновым дыханием.
Не у всех растений фотодыхание в равной степени происходит интенсивно. У сои оно идет с большей скоростью, а у сахарного тростника его долгое время не могли выявить. Вообще, у С3-растений наблюдается более интенсивное фотодыхание, чем в С4-растениях.
Интенсивное фотодыхание может снизить продуктивность С3-растений на 30 %, так как при фотодыхании происходит не поглощение СО2, а его потери. Из каждых 10 молекул рибулозо-1,5-дифосфата, образовавшихся в С3-цикле и включившихся в фотодыхание, одна разрушается.
В связи с этим важной задачей для физиологов и селекционеров растений считается выведение сортов растений, у которых в максимальной степени проявляется карбоксилазная и подавлена оксигеназная реакция и, таким образом, подавлено фотодыхание.
Низкую интенсивность фотодыхания в С4-растениях объясняют тем, что в них высокая концентрация СО2 в клетках-обкладках. В этих растениях РДФ-карбоксилаза находится только в клетках-обкладках, где и происходит процесс. Некоторые исследователи считают, что в С4-растениях фотодыхание подавлено
из-за недостатка О2 в клетках-обкладках. В этих клетках не идет фотолиз воды, так как они не имеют ФС II, а поступление атмосферного кислорода затруднено, потому что эти клетки расположены в глубине листовой пластинки и между ними почти нет межклетников.
В настоящее время фотодыхание рассматривают как одну из необходимых функций зеленого растения. Во-первых, фотодыхание, как и С3-цикл, CAM-цикл и другие процессы, является источником метаболитов для разных синтезов. Во-вторых, практически доказано, что при образовании серина и глицина восстанавливается митохондриальная НАД+. В-третьих, выявлена связь фотодыхания с ассимиляцией азота. И, наконец, появилась теория, в соответствии с которой излишки продуктов цикла Кальвина служат сигналом, который увеличивает фотосинтез. Таким образом, соотношение фотосинтеза и фотодыхания выполняет регуляторную роль в растениях.
|
|
|
