 |


Конструктивный метаболизм бактерий
|
|
|
|
Конструктивный метаболизм бактерий
Основные компоненты клетки синтезируются из аминокислот, фосфатов, пуриновых и пиримидиновых оснований, органических кислот. Поставщиками этих веществ являются в том числе и промежуточные продукты энергетического метаболизма.
Выше было сказано, что среди бактерий выделяют прототрофы, которые способны синтезировать все компоненты клетки из одного источника углерода и энергии. В случае, когда бактерии теряют способность синтезировать какой-либо фермент, участвующий в биосинтетических реакциях, то для их роста и размножения требуется недостающее вещество, которое называется фактором роста. Такие бактерии называются ауксотрофами. Факторами роста могут быть аминокислоты, пуриновые и пиримидиновые основания, витамины.
Биосинтез белков. В процессе катаболизма бактерии разлагают белки под действием протеаз с образованием пептидов. В последующем под действием пептидаз пептиды разрушаются до аминокислот. В результате распада аминокислот клетка получает ионы аммония, которые требуются для синтеза собственных аминокислот. Большинство бактерий обладает способностью синтезировать все 20 аминокислот, из которых состоят белки. Белки в бактериальной клетке выполняют ферментативную функцию, а также являются составной частью структурных компонентов клетки (ЦПМ, ее производных, клеточной стенки, жгутиков, капсулы, спор).
Углеродные скелеты аминокислот образуются из промежуточных продуктов обмена. Исходным материалом служат промежуточные продукты фруктозодифосфатного (ФДФ) и пентозофосфатного (ПФ) путей, цикл трикарбоновых кислот: пируват, кетоглутаровая кислота, оксалоацетат, фумарат, эритрозо-4-фосфат, рибозо-4-фосфат. Аминогруппы в состав аминокислот вводятся в
|
|
|
результате непосредственного аминирования или переаминирования. Перевод неорганического азота в органические соединения происходит всегда через аммиак. Нитраты, нитриты и молекулярный азот предварительно восстанавливаются в аммиак и только лишь после этого включаются в состав органических соединений.
В результате прямого аминирования образуются лишь L-аланин, L-аспартат, L-глутамат и L-глутамин. Все остальные аминокислоты получают свою аминогруппу в результате переаминирования, с одной из “первичных” аминокислот. В большинстве случаев аминогруппа вводится на последнем этапе синтеза путем переаминирования.
Биосинтез нуклеотидов. Пуриновые и пиримидиновые нуклеотиды – это те строительные блоки, из которых синтезируются нуклеиновые кислоты. Кроме того, пуриновые и пиримидиновые нуклеотиды входят в состав многих коферментов и служат для активации и переноса аминокислот, сахаров, липидов в реакциях полимеризации.
Исходным соединением для образования пентозной части нуклеотидов служит рибозо-5-фосфат, образующийся в ПФ-пути. Углеродный скелет пиримидинов происходит из аспартата, который образуется в цикле трикарбоновых кислот. Атомы азота и аминогруппы пуринов и аминосодержащих пиримидинов происходят из аспартата и глутамина.
Биосинтез липидов. Жиры или липиды являются важными компонентами ЦПМ и клеточной стенки грамотрицательных бактерий, а также служат запасными веществами. В бактериальных жирах преобладают длинноцепочечные (С14- - С18) насыщенные жирные кислоты и ненасыщенные жирные кислоты, содержащие одну двойную связь. Сложные липиды представлены фосфатидилинозитом, фосфатидилглицерином и фосфатидилэтаноламином.
Ключевым промежуточным продуктом для биосинтеза жирных кислот является ацетил-коэнзим А. Ключевыми промежуточными продуктами для синтеза фосфолипидов является продукт ФДФ-пути: диоксиацетилфосфат, восстанавливающийся в глицеро-3-фосфат, который соединяется с остатками жирных кислот.
|
|
|
Биосинтез углеводов. Углеводы представлены в бактериальной клетке в виде моно-, ди- и полисахаридов, а также комплексных соединений. Полисахариды входят в состав некоторых капсул, крахмал и гликоген являются запасными питательными веществами.
Синтез глюкозы происходит из пирувата, за счет обратных реакций, путей распада глюкозы. Для обхода реакций, идущих только в одном направлении, имеются обходные пути, например, глиоксилатный цикл.
Транспорт веществ из бактериальной клетки
Для обеспечения жизнедеятельности бактерии должны не только поглощать из внешней среды питательные вещества, но и выделять в окружающую среду гидролитические ферменты, токсины и другие соединения (метаболиты).
У грамположительных микробов белки секретируются непосредственно во внешнюю среду. У грамотрицательных бактерий секретируемые соединения
должны транспортироваться через две мембраны клеточной оболочки. Наличие наружной мембраны привело к тому, что у грамотрицательных бактерий в процессе эволюции сформировалось несколько систем секреции, различающихся по структуре и функциям. В настоящее время у бактерий выявлено 7 систем секреции.
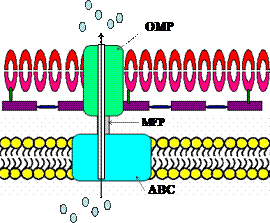
Белки, секретируемые по I пути ( система секреции I типа, T1SS), пересекают внутреннюю цитоплазматическую и наружную мембраны в один этап в результате образования во внутренней мембране, периплазматическом пространстве и наружной мембране одной поры (канала). Эта система секреции включает 3 компонента: АТФ-связывающую кассету (АВС-компонент, ATP-binding cassette), белки, соединяющие мембраны (MFP, membrane fusion protein) и белки наружной мембраны (OMP, outer membrane protein). Секреция в этом случае протекает без участия sec-белков (рисунок 27).
 Наружная мембрана
Наружная мембрана
|
|
|
