 |


Рост и размножение бактерий
|
|
|
|
ЦПМ
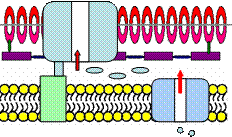
Рисунок 27 – Бактериальная система секреции I типа.
Этим путем секретируются пороформирующие токсины: гемолизины, металлопротеазы, некоторые гидролитические ферменты, внеклеточная аденилатциклаза B. pertussis.
Система секреции II типа (T2SS) обеспечивает секрецию гидролитических ферментов, некоторых токсинов, протеинов, участвующих в формировании поверхностных структур бактериальной клетки (в частности, пилей). Эта система секреции широко представлена среди грамотрицательных бактерий, в связи с чем ее называют общим секреторным путем (GSP – general secretory pathway). Белки, секретируемые по II пути, проходят через внутреннюю и наружную мембраны отдельными этапами при участии sec-белков. После транслокации через внутреннюю мембрану секретируемые белки задерживаются в периплазматическом пространстве, где изменяют свою конформацию. В результате таких изменений секретируемый белок приобретает окончательную структуру и транспортируется через наружную мембрану (рисунок 28).


 Наружная мембрана
Наружная мембрана
ЦПМ
 | |||
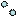 | |||
Рисунок 28 - Бактериальная система секреции II типа.
По II типу из бактериальных клеток выделяется холерный токсин, фосфолипаза С, эластаза, экзотоксин А и другие факторы патогенности P. aeruginosa.
Система секреции III типа (T3SS) грамотрицательных бактерий в основном предназначена для транспорта из клетки компонентов жгутиков. Этот путь используется также для направленной доставки в клетку хозяина бактериальных белков - эффекторов. В результате действия этих белков нарушаются функции клетки хозяина. Система секреции III типа представляет собой шприцеподобную структуру (инъектисому), состоящую из нескольких белков и способную инъецировать эффекторные белковые молекулы непосредственно в цитозоль клетки хозяина (рисунок 29).
|
|
|
 Наружная мембрана
Наружная мембрана
ЦПМ
Рисунок 29 - Бактериальная система секреции III типа.
Эффекторные белки вызывают реорганизацию цитоскелета клетки хозяина, в результате чего бактерии проникают в клетку. Система секреции III типа обнаружена у Salmonella, Shigella, Yersinia, P. aeruginosa, Chlamydia, некоторых энтеропатогенных E. coli, других бактерий. T3SS вносит существенный вклад в развитие патогенеза заболевания. Строение системы секреции III типа сальмонелл, полученное с помощью электронного микроскопа, представлено на рисунке 30.

Рисунок 30 - Строение системы секреции III типа. А – электронная
микрофотография зоны контакта бактериальной клетки и клетки хозяина; В – инъектисомы в свободном состоянии; С – схема строения инъектисомы сальмонелл
(обозначены белки, формирующие аппарат секреции).
Белки, секретируемые по системе III типа, пересекают внутреннюю цитоплазматическую и наружные мембраны в один этап без участия sec-белков.
Система секреции IV типа (T4SS) Белки, секретируемые по IV пути, проходят через внутреннюю и наружную мембрану отдельными этапами при участии sec-белков (рисунок 31).
 Наружная мембрана
Наружная мембрана
ЦПМ
 |
Рисунок 31 - Бактериальная система секреции IV типа.
Система секреции IV типа встречается у Helicobacter pylori, легионелл, бруцелл и других патогенов. В частности, с помощью этой системы бруцеллы после фагоцитоза вводят через стенку фагосомы в цитоплазму эукариотической клетки эффекторные белки, препятствующие слиянию фагосомы с лизосомой.
Система секреции V типа (T5SS) отличается тем, что в периплазматическом пространстве из части секретируемого полипептида формируется цилиндрическая структура, выполняющая роль поры, через которую белок выходит наружу. По этому пути секретируются IgA-протеаза у N. gonorrhoeae, белок пертактин у B. pertussis, адгезины многих патогенов (рисунок 32).
|
|
|

 Наружная мембрана
Наружная мембрана
ЦПМ
 |  |


Рисунок 32 - Бактериальная система секреции V типа.
Система секреции V типа называется также системой автотранспортеров, так как в ней С-концевая последовательность белка отвечает за экспорт N- терминального домена через наружную мембрану. С помощью этой системы секреции транспортируются белки большой молекулярной массы, состоящие не менее чем из 3000 аминокислотных остатков.
Система секреции VI типа (T6SS) обеспечивает доставку секретируемых белков непосредственно в цитоплазму клеток хозяина. Эта система секреции встречается у Vibrio cholerae, Pseudomonas aeruginosa, Francisella tularensis и других патогенов. T6SS представляет собой полую структуру, состоящую из колец. В процессе сборки она постепенно удлиняется, проходит через наружную мембрану, растет до соприкосновения с мембраной эукариотической клетки, прокалывает ее и осуществляет инъекцию эффекторных молекул (рисунок 33).
 Наружная мембрана
Наружная мембрана
ЦПМ
Рисунок 33 – Бактериальная система секреции VI типа.
Система секреции VII типа (T7SS) характерна для микобактерий (Mycobacterium tuberculosis, M. bovis, M. avium, M. marinum). В составе этой системы секреции присутствует две поры. Одна пора располагается в цитоплазматической мембране, а другая - в микомембране - наружной части клеточной стенки, содержащей липиды (рисунок 8. 34).
 Микомембрана
Микомембрана
ЦПМ
Рисунок 34 – Бактериальная система секреции VII типа.
Таким образом, среди бактериальных систем секреции описаны как одноэтапные (одношаговые), так и двухэтапные (двухшаговые). У одноэтапных систем секреции пренос белков происходит непосредственно из цитоплазмы во внешнюю среду. У двухэтапных систем секреции секретируемый белок вначале переносится из цитоплазмы бактериальной клетки в периплазматическое пространство, а затем – во внешнюю среду.
Рост и размножение бактерий
Полученная клеткой энергия в процессе дыхания или брожения используется для роста и размножения бактерий.
Рост - это увеличение массы отдельной клетки в результате синтеза клеточного материала. Во время роста размеры клетки увеличиваются. После достижения определенных размеров клетка прекращает рост и начинает делиться (размножаться). Размножение бактерий – это способность бактерий к самовоспроизведению (увеличению количества особей).
|
|
|
 |  | ||
Для подавляющего большинства бактерий характерно бинарное поперечное деление, приводящее к образованию двух одинаковых дочерних клеток (рисунок 35).
а б
Рисунок 35 - Бинарное деление бактериальной клетки: а – электронная микрофотография; б – схема деления.
Деление бактериальной клетки начинается после завершения репликации бактериальной хромосомы. При репликации каждая из двух нитей ДНК служит матрицей для синтеза комплементарной дочерней цепи ДНК. В репликации участвует фермент ДНК-полимераза. В результате удвоения родительской ДНК происходит образование двуспиральных дочерних молекул ДНК. Затем в средней части клетки формируется поперечная перегородка (септа), которая делит цитоплазму материнской клетки на две дочерние. Одновременно с этим синтезируется клеточная стенка, образующая полноценную перегородку между двумя дочерними клетками.
Некоторые микроорганизмы размножаются путем почкования. Особенно этот способ характерен для дрожжей. При почковании на одном из полюсов материнской клетки образуется маленький вырост (почка). Постепенно почка достигает размеров материнской клетки и отделяется от нее (рисунок 36).
 | 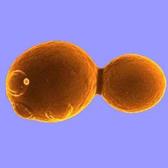 | ||
Рисунок 36 - Почкование дрожжей.
Размножение бактерий изучают при их культивировании в жидкой или на плотной питательных средах. В жидкой питательной среде культивирование может носить непрерывный или периодический характер. Для производства биомассы бактерий применяют культивирование в аппаратах-культиваторах (рисунок 37).
 |
Рисунок 37 – Схема аппарата-культиватора.
При непрерывном культивировании используется открытая система с возможностью подачи свежей питательной среды и вывода из аппарата- культиватора (ферментера) накопленной биомассы или метаболитов. При
|
|
|
периодическом культивировании питательные вещества в систему дополнительно не вводятся, а продукты обмена не удаляются. В этом случае размножение бактерий в жидкой питательной среде характеризуется сменой фаз или стадий (рисунок 38):
- лаг-фаза - начальная стадия адаптации бактерий к питательной среде. В эту фазу наблюдается синтез адаптивных ферментов. Лаг-фаза представляет собой период от момента внесения (посева) бактерий в питательную среду до их роста. В этой фазе число живых бактерий не увеличивается, а может даже уменьшаться. Продолжительность этой фазы составляет 2-4 часа;
- экспоненциальная (логарифмическая) фаза (лог-фаза) – фаза выраженного прироста численности бактерий (увеличение количества микробных клеток в геометрической прогрессии: в конце первой генерации из одной клетки образуется две, в конце второй - четыре и так далее). В этот период скорость размножения клеток и увеличение бактериальной популяции максимальны. Период генерации, то есть период времени между двумя последовательными делениями бактерий, постоянный для данного вида микроорганизмов. Продолжительность этой фазы - 5-6 часов;
- стационарная фаза - фаза максимального накопления клеток и развития равновесия между размножением и гибелью клеток. Число новых бактерий почти равно числу отмерших, то есть наступает равновесие между погибшими клетками и вновь образующимися клетками. Увеличения численности микроорганизмов в эту стадию не происходит. Продолжительность этой фазы - 2 часа;
- фаза отмирания - уменьшение численности популяции в связи с уменьшением питательных веществ и изменением условий культивирования (изменение рН, концентрации ионов и др. ). Продолжительность этой фазы - около 5 часов. Остающиеся в живых клетки переходят в состояние покоя.
 |
Рисунок 38 – Размножение бактерий в жидкой питательной среде.
Большинство микроорганизмов при благоприятных условиях делятся пополам через каждые 20-30 минут. При такой скорости деления из одного микроорганизма через 5 часов образуется 1024 особей.
Размножение бактерий происходит на питательных средах, содержащих необходимые компоненты, в оптимальных температурных условиях, при определенном насыщении среды кислородом.
Размножение характеризуется:
- временем генерации - интервалом времени, за который число клеток удваивается;
- концентрацией бактерий - числом клеток в 1 мл.
|
|
|
По температурному оптимуму роста выделяют три основные группы микроорганизмов.
1. Психрофилы - растут при температурах ниже +20ОС.
2. Мезофилы - растут в диапазоне температур от +20ОС до +45ОС (в основном оптимальная температура составляет 37ОС).
3. Термофилы - растут при температурах выше +45ОС.
|
|
|
