 |


1.4.3 Катаболизм серотонина. 1.4.4 Рецепторы к серотонину
|
|
|
|
1. 4. 3 Катаболизм серотонина
Серотонин, захваченный из синаптической щели назад в нейрон или глиальную клетку, разрушается с помощью фермента моноаминоксидазы типа А (МАО-А) (рисунок 6), дезаминирующая активность которого у больных психическими расстройствами повышена по сравнению с нормой серотонина (Максимова с соавт., 2007).
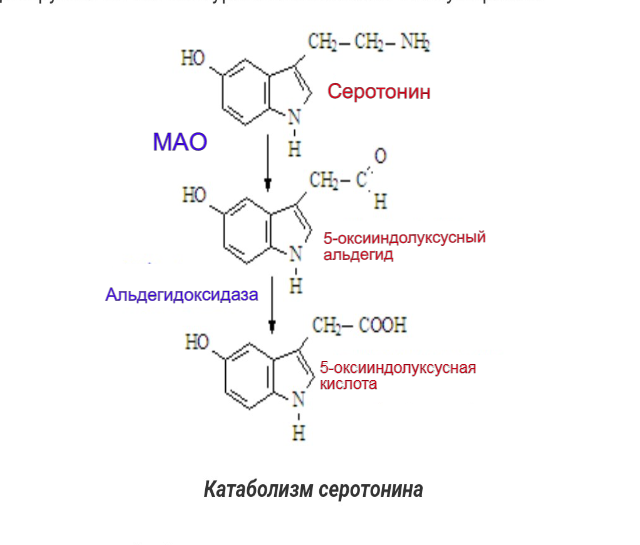
Рисунок 6– Катаболизм серотонина (Рычков, 2014)
МАО - это митохондриальные ферменты, содержащие флавин, катализирующие окислительное дезаминирование нейромедиаторов и биогенных амидов в головном мозге и периферических тканях. На основании селективности к субстрату и селективности ингибитора были обозначены две формы МАО: МАОА и МАОБ, которые соответствуют двум различным генам. Обычно MAOA катализирует окисление серотонина (5-Ht), тогда как MAOБ действует на 2-фенилэтиламин и бензиламин. Дофамин, норадреналин, адреналин, триптамин и тирамин окисляются обеими формами фермента у большинства видов (Youdim, Bakhle, 2006), тогда как норадренергические нейротрансмиттеры предпочтительно дезаминируются ферментами МАОА. По своему строению эти белки сходны между собой, их аминокислотные последовательности совпадают примерно на 70 %. Т. е. это ферменты, осуществляющие катаболизм моноаминов посредством их окислительного дезаминирования. МАО метаболизирует как эндогенные моноамины — нейромедиаторы и гормоны, так и экзогенные — попадающие в организм с пищей или в лекарствах и психоактивных веществах. Исследования показали, что активность МАО определяется генетически и не изменяется в течение жизни в физиологических условиях. MAO-A и Б кодируются отдельными генами, расположенными на Х-хромосоме. Каждый ген состоит из 15 экзонов с разными коровыми промоторными областями, но с идентичной организацией интрон-экзон, что указывает на то, что MAO-A и Б происходят от дупликации общего предкового гена.
|
|
|
Метаболизм 5-Ht происходит под действием моноаминоксидазы A (MAO-A) с образованием метаболита 5-гидроксииндолуксусной кислоты (5-HIAA) (рисунок 7). Поскольку MAO-A является внутриклеточным ферментом, 5-Ht должен быть поглощен внутри клетки до того, как на него будут воздействовать, и как Sert, так и транспортер норадреналина (Net) способствуют этому поглощению (Stoltenberg et al., 2002). Ткани или клетки, которые вносят значительный вклад в метаболизм 5-Ht, включают клетки легких, кишечника и эндотелия артериальной системы, но любая клетка, которая может принимать 5-Ht и обладает MAO-A, может метаболизировать 5-Ht.

Рисунок 7– Серотонинергический синапс и обращение с серотонином (5-Ht) в результате синтеза, хранения, высвобождения, поглощения и метаболизма (Palczewski, Kiser 2013)
1. 4. 4 Рецепторы к серотонину
Поразительная полифункциональность серотонина обусловлена, прежде всего, наличием многочисленных рецепторов, опосредующих воздействие этого медиатора на нейроны (Попова, Науменко, 2010). К настоящему времени с помощью молекулярно-биологических методов выявлено и клонировано 14 различных типов генов и контролируемых ими серотонергических (5-Нt) рецепторов, экспрессирующихся в мозге млекопитающих.
Классификация 5-Нt рецепторов основана на трех принципах:
а) особенностях первичной структуры;
б) механизмах трансдукции сигнала;
в) на фармакологическом профиле (селективные агонисты и антагонисты рецептора).
По этим принципам 5-Нt рецепторы классифицированы в 8 основных типов и 7 подтипов. Большинство 5-Нt рецепторов принадлежат к суперсемейству рецепторов, сопряженных с G-белками (таблица 1).
Каждый из них представляет собой полипептидную цепь, содержащую семь трансмембранных доменов, образующих совместно с внеклеточными петлями и N-концом участок связывания медиатора. Третья цитоплазматическая петля и С-конец осуществляют сопряжение с G-белком. При связывании медиатора происходит изменение конформации рецептора, приводящее к диссоциации G-белка на β -, γ - и α -субъединицы. Эти субъединицы обладают каталитической активностью, благодаря которой способны изменять внутриклеточные процессы.
|
|
|
Таблица 1 - Общая характеристика типов серотониновых рецепторов. (Qi et al., 2014)
| Тип | Механизм работы | Эффекты при активации | Генерирующий потенциал |
| 5-Ht1 | Gi/G0-связанный | ↓ yровня цАМФ | Тормозящий |
| 5-Ht2 | Gq/G11-связанный | ↑ уровня инозитол-3-фосфата (IP3) и диацилглицерола (DAG) | Возбуждающий |
| 5-Ht3 | Лиганд-зависимый ионный канал, пропускающий натрий и калий | Деполяризация клеточной мембраны | Возбуждающий |
| 5-Ht4 | Gs-связанный | ↑ уровня цАМФ | Возбуждающий |
| 5-Ht5 | Gi/G0-связанный | ↓ уровня цАМФ | Тормозящий |
| 5-Ht6 | Gs-связанный | ↑ уровня цАМФ | Возбуждающий |
| 5-Ht7 | Gs-связанный | ↑ уровня цАМФ | Возбуждающий |
| 5-Ht8 | Метаботропный неуточнённый | ↑ внутриклеточного уровня кальция | Неизвестно |
Каждый из типов и подтипов рецепторов характеризуется уникальным контролирующим геном, спектром сродства к разнообразным агонистам и антагонистам серотонина, специфическим распределением в мозге и определенным спектром функций, которые он регулирует. Среди такого разнообразия серотониновых рецепторов привлекает внимание 5-Нt1а подтип. Это обусловлено рядом причин, основными из которых являются:
1) ключевая роль в ауторегуляции 5-Ht системы в мозге (Popova, Naumenko, 2013). 5-Нt1а рецепторы отличаются от других видов 5-Нt рецепторов тем, что они локализованы как пост-, так и пресинаптически на 5-Нt нейронах, и в зависимости от локализации могут оказывать прямо противоположное действие;
2) экспериментальные данные о вовлечении 5-Нt1а рецепторов в регуляцию разнообразных форм как нормального, так и патологического поведения;
3) многочисленные данные об участии 5-Нt1а рецепторов в механизме патологической тревожности и депрессии (Kaufman et al., 2015), подтвержденные клинической эффективностью агонистов 5-Нt1а рецепторов как анксиолитиков и антидепрессантов;
|
|
|
4) наличие селективных агонистов и антагонистов 5-Нt1а рецепторов. Из-за многообразия функций 5-Нt1а рецепторов встает вопрос об их генетической и посттранляционной регуляции, связи предрасположенности к патологическому поведению с особенностями генетического контроля, плотности и функциональной активности этих рецепторов. В этом обзоре будут обсуждены данные литературы и собственных исследований, касающиеся генетических подходов к исследованию 5-Нt1а рецепторов и их участия в регуляции поведения.
Рецепторы типа 5-НТ1
В центральной нервной системе (ЦНС) млекопитающих обнаружены серотониновые рецепторы типа 5-Ht1 и пять их подтипов – a, b, d, e, f, представляющие собой протеины, содержащие 365-422 аминокислотных остатка. Эти рецепторы связаны с ингибиторным G-белком, подавляющим аденилатциклазу, за счёт чего подавляется активность внутриклеточных процессов.
5-Нt1а серотониновый рецептор 5-Нt1а рецептор является эволюционно одним из самых древних. Считается, что он образовался около 800 млн. лет назад, задолго до появления позвоночных животных. Он обнаружен у насекомых (Dacks et al., 2006), плоских червей, круглоротых (Hill et al., 2003) и всех классов позвоночных животных. 5-Нt1а подтип относится к группе рецепторов 5-НТ1 типа, которая также включает в себя 5-Нt1а, 5-Ht1b, 5-Ht1d, 5-Ht1e и 5-Ht1f подтипы, проявляющие от 40 до 63 % структурной гомологии с 5-Нt1а рецептором. Рецепторы 5-Нt1 типа сопряжены с Gi-белком и при активации ингибируют аденилатциклазу – фермент, катализирующий превращение АТФ в цАМФ, что является сигналом для активации цАМФзависимых протеинкиназ, которые в свою очередь активируют различные белки (в том числе факторы транскрипции). Механизм работы 5-Ht1a рецептора изображен на рисунке 8. Активация 5-Нt1 рецепторов приводит к открытию К+ каналов и гиперполяризации мембраны. Имеются данные о том, что активация рецептора 1А подтипа в культурах клеток может приводить к активации фосфолипазы С.

Риcунок 8– Механизм работы 5-Ht1a рецептора (Бохан с соавт. 2013)
|
|
|
5-Нt1а рецептор представляет собой полипептидную цепь, образующую семь трансмембранных доменов и состоящую из 422 аминокислотных остатков у крысы и 421 – у человека. Этот рецептор характеризуется коротким внутриклеточным С-концом и длинной третьей внутриклеточной петлей, на которой локализован сайт фосфорилирования протеинкиназой С. Была показана связь между фосфорилированием и десенситизацией рецептора, что дало основание полагать, что функция 5-Нt1а рецепторов находится под контролем 5-Нt2 рецепторов, вторичным посредником которых является протеинкиназа С (Zhang et al., 2001). Кроме того, в третьей цитоплазматической петле имеются два сайта связывания с кальмодулином, связывание которого является сигналом для интернализации рецептора (Turner et al., 2004). Еще одной особенностью структуры рецептора А подтипа является наличие «лейциновых застежек» (leucine zipper), посредством которых, возможно, осуществляются белок-белковые взаимодействия между 5-Нt1а и 5-Нt2 рецепторами. Все эти особенности структуры 5-Нt1а рецептора приоткрывают возможные пути воздействия на его экспрессию, в том числе другими 5-Нt рецепторами. Факторы, влияющие на экспрессию рецептора, могут, по-видимому, действовать и на уровне транскрипции. В 1999 году была описана структура промотора гена 5-Нt1а рецептора. Было показано, что основной сайт инициации транскрипции локализован в положении –967 п. н. от стартового кодона AUG. У крыс этот сайт ассоциирован с выше расположенными TATA-боксами, однако у мышей TATA-боксы отсутствуют. Принимая во внимание сходство этих животных в экспрессии рецептора и ее регуляции, авторы предположили, что TATA-боксы не играют существенной роли в регуляции транскрипции гена 5-Нt1а рецептора. Кроме того, в промоторе было обнаружено наличие селективного энхансера (enhancer) (между –426 и –117 п. н. ), неселективного (между –1519 и –426 п. н. ) и сильного селективного сайленсера (silencer) (между –1590 и –1519 п. н. ) (Ou et al., 2000). Из этих элементов наиболее интересным является сайленсер, поскольку в случае его делеции транскрипция гена усиливается более чем в 10 раз. Поэтому было предположено, что регуляция экспрессии гена 5-Нt1а рецептора на уровне транскрипции осуществляется, вероятно, в основном за счет репрессии (Ou et al., 2001). Таким образом, наряду с генетическим контролем интенсивности синтеза 5-Ht1а рецепторов существуют и другие уровни регуляции их функциональной активности.
В последние годы к выявленным ранее механизмам регуляции 5-Нt1а рецептора добавлены новые, связанные с олигомеризацией и взаимодействием 5-Нt рецепторов. Было установлено, что 5-Нt1а рецептор существует в мозге в нескольких формах: как 5-Нt1а мономер, 5-Нt1а/5-Нt1а гомодимер и гетеродимер, связанный с другими рецепторами (Woehler et al., 2009; Kobe et al., 2012; Renner et al., 2012).
|
|
|
Новые данные о роли димеризации в регуляции функциональной активности 5-Нt1а рецептора получены при изучении взаимодействия 5-Нt1а и 5-Нt7 рецепторов (Renner et al., 2012; Popova, Naumenko, 2013; Naumenko et al., 2014). Оказалось, что гетеродимер 5-Нt1а/5-Нt7, не оказывая влияния на функциональные свойства 5-Нt7 рецептора, модулирует два основных фактора десенситизации 5-Нt1а рецепторов: ослабляет их функциональную активность и увеличивает интенсивность интернализации. Таким образом, ключевой регулятор 5-Нt системы мозга, 5-Нt1а рецептор, сам зависит от 5-Нt7 рецепторов, которые, образуя с ним димерные комплексы, ведут к функциональной инактивации. Это совершенно новая и важнейшая роль 5-Нt7 рецептора и его взаимодействия (cross-talk) с 5-Нt1a рецептором позволяет по-новому взглянуть на проблему механизма депрессии и действия антидепрессантов группы ингибиторов обратного захвата серотонина.
Рецепторы типа 5-Нt2
В отличие от первого типа, они являются возбуждающими. Рецепторы 5-Нt2 увеличивают чувствительность нейрональной мембраны к стимулам. Они уменьшают поступление ионов калия (К+) в клетку и облегчают посредством данного эффекта возникновение деполяризации, в этом отношении они оказывают эффект, противоположный 5-Нt1-рецепторам. Механизм работы 5-Нt2а-рецептора изображен на рисунке 9.

Рисунок 9– Механизм работы 5-Нt2а-рецептора (Palczewski, Kiser 2013)
При активации рецептора бета- и гамма-субъединицы «высвобождают» Gq-субъединицу, активирующую фосфолипазу С (PLC), которая, в свою очередь, превращает фосфатидилинозитолбисфосфат (PIP2) в диацилглицерол (DAG) и инозитолтрифосфат (IP3). DAG активирует протеинкиназу С (PKC), а IP3 запускает кальмодулин-зависимый механизм высвобождения кальция из эндоплазматического ретикулума.
Существуют также побочные биохимические пути, связанные с образованием арахидоновой кислоты из DAG. «Выбор» пути, по которому будут работать вторичные мессенджеры, может также зависеть от лиганда: например, если рассматривать DOB и 2C-B, то DOB (за счёт наличия метильной группы в альфа-положении на боковой цепи) действует по «основному» пути, описанному выше, а 2C-B – по «альтернативному», ведущему к повышению концентрации арахидоновой кислоты.
5-Нt3-рецептор.
Данный вид рецепторов является уникальным в своём роде – это единственный неметаботропный тип. 5-Нt3-рецептор представляет собой лиганд-зависимый ионный канал, состоящий из пяти различных субъединиц (5-Ht3a, 5-Ht3в, 5-Ht3с, 5-Ht3d, 5-Ht3e). Он может быть представлен в виде гомопентамера, состоящего исключительно из одних 5-Нt3а-субъединиц, либо гетеропентамера – когда в его составе, помимо обязательно наличествующей 5-Нt3а-субъединицы, находятся субъединицы других подтипов. Интересно, что по уровню гомологии субъединицы можно разделить на 2 группы: первая включает в себя 5-Ht3a и 5-Ht3в, вторая 5-Ht3с, 5-Ht3d и 5-Ht3e.
Каждая субъединица состоит из четырёх участков(М1-М4), представляющих собой аминокислотную цепь, уложенную в форме альфа-спиралей, которые пересекают клеточную мембрану. N-конец аминокислотной цепи субъединицы локализован на внешней стороне клеточной мембраны и осуществляет связывание лигандов, трансмембранные участки (а конкретнее – аминокислотная петля, связывающая М2 и М3-участки) непосредственно задействованы в механизме открытия ионного канала. Также присутствует внутриклеточная петля между М3 и М4-участками, участвующая в транспортировке и регуляции работы субъединицы. Механизм работы 5-Нt3-рецептора изображен на рисунке 10.

Рисунок 10– Механизм работы 5-Нt3-рецептора (Naumenko, 2013)
Данный ионный канал пропускает только положительно заряженные ионы (натрий, калий, небольшие количества кальция), что вызывает деполяризацию и возбуждение нейрона при его активации.
5-Ht3 рецептор оказывает стимулирующее влияние на функцию GABA-ергических нейронов в гиппокампе и новой коре головного мозга. Потому что GABA является ингибирующим нейротрансмиттером, что приводит в итоге к уменьшению его активности. Нейрофизиологические свойства указывают на то, что сам 5-Нt3-рецептор не является G-белок-связанным рецептором. Активность ионного канала регулируется ионами кальция и магния (Ca2+ и Mg 2+).
5-Нt4-рецептор, вероятно, является родственным 5-Нt1-типу. Через влияние на транспорт ионов 5-Нt4-рецепторы ингибируют гиперполяризацию нейронов и вследствие этого увеличивают их чувствительность. Периферические и центрально расположенные 5-Нt4-рецепторы играют значимую роль как пресинаптические гетерорецепторы на холинергических окончаниях нервных клеток. Стимуляция этих нейронов приводит к облегчению выделения ацетилхолина. Данный вид рецепторов является метаботропным, при активации повышающим количество цАМФ в клетке, что в свою очередь запускает каскад реакций, ведущий к выбросу нейромедиаторов. Ранее он считался «кардиоспецифичным» рецептором, так как изначально был обнаружен в наибольших концентрациях в предсердных отделах проводящей системы сердца.
Вклад мутаций гена этого рецептора в развитие шизофрении достаточно противоречив, однако их связь с возникновением биполярного расстройства подтверждёна достоверно.
5-Нt5 и 5-Нt6-рецепторы
На сегодняшний день всё ещё немногое известно о функции 5-Нt5 и 5-Нt6-рецепторов. Установлено, что оба рецептора стимулируют активность аденилатциклазы.
Среди всех рецепторов серотонина тип 5-Ht5 является наименее изученным. Пока известно лишь то, что в геноме человека представлено 2 подтипа – 5-Нt5а и 5-Нt5в. Однако, ген второго подтипа является псевдогеном, т. к. в его последовательности содержится стоп-кодон, что делает его нефункциональным. У грызунов гены обоих подтипов рецептора являются функциональными.
5-Нt7-рецепторы
5-Нt7-рецептор осуществляет свое действие через влияние на активность аденилатциклазы. Эти метаботропные серотониновые рецепторы также запускают процессы высвобождения нейромедиаторов путём запуска каскада реакций фосфорилирования через цепь вторичных мессенджеров, т. е. являются «возбуждающими». Изначально они были идентифицированы в качестве подтипа 5-Нt1-рецепторов, однако позже, благодаря изучению их структуры и аминокислотной последовательности, были выделены в отдельный тип. Этот рецептор играет очень большую роль в определении биоритма. Новые данные о роли димеризации в регуляции функциональной активности 5-Нt1а рецептора получены при изучении взаимодействия 5-Нt1а и 5-Нt7 рецепторов (Renner et al., 2012; Popova, Naumenko, 2013; Naumenko et al., 2014). Оказалось, что гетеродимер 5-Нt1а/5-Нt7, не оказывая влияния на функциональные свойства 5-Нt7 рецептора, модулирует два основных фактора десенситизации 5-Нt1а рецепторов: ослабляет их функциональную активность и увеличивает интенсивность интернализации. Таким образом, ключевой регулятор 5-Нt системы мозга, 5-Нt1а рецептор, сам зависит от 5-Нt7 рецепторов, которые, образуя с ним димерные комплексы, ведут к функциональной инактивации. Это совершенно новая и важнейшая роль 5-Нt7 рецептора и его взаимодействия (cross-talk) с 5-Нт1a рецептором позволяет по-новому взглянуть на проблему механизма депрессии и действия антидепрессантов группы ингибиторов обратного захвата серотонина.
|
|
|
