 |


Симуляция читающей коры. Предпочтения нейронов и их роль в чтении
|
|
|
|
Симуляция читающей коры
По всей вероятности, точное представление о нейронной архитектуре для чтения можно получить только одним способом: смоделировать формирование этого навыка на компьютере, строение которого напоминает структуру коры. Однако в настоящее время такая симуляция остается недостижимой мечтой. Чтобы воспроизвести воздействие тысяч часов чтения на миллионы нейронов в области «буквенной кассы», требуется суперкомпьютер, гораздо более мощный, чем любая из машин, существующих на сегодняшний день. Пока мы можем только надеяться, что прогресс в области вычислительной техники, наряду с появлением новых компьютеров, предназначенных для моделирования нейронных сетей, в результате позволит создать адекватную модель читающего мозга.
Тем временем стоит задуматься о том, как такая симуляция могла бы объединить огромное количество существующих систем письменности. В принципе архитектура мозга, читающего латинский алфавит, должна быть применима и к китайским иероглифам. Под влиянием китайского языка в нем должны развиться нейроны, настроенные на часто встречающиеся символы и их внутренние компоненты. В самом деле, большинство иероглифов содержат ограниченное количество семантических и фонетических маркеров, состоящих из нескольких стандартизированных штрихов – встроенного набора зрительных закономерностей, которые легко может уловить иерархический алгоритм обучения. Поведенческие эксперименты показывают, что чтение китайских символов основывается именно на такой иерархической и комбинаторной схеме[269]. Что касается китайских аналогов биграммных нейронов, то пока они не открыты.
|
|
|
Вероятно, что даже у читателей латинского алфавита нейроны со временем становятся чувствительны к единицам разного размера. Для языков с прозрачной орфографией, например итальянского, достаточно относительно скромного запаса детекторов букв и биграмм. Языки с непрозрачной орфографией, такие как английский или французский, напротив, требуют гораздо большего количества нейронов для кодирования таких больших и сложных орфографических единиц, как «ough», «tion» или «ould»[270]. Подобные зрительные закономерности, выходящие за пределы уровня букв, могут быть извлечены только на высших уровнях зрительной иерархии. Таким образом, наша модель предсказывает, что чтение на английском языке, в отличие от чтения на итальянском, должно задействовать области не только более обширные, но и расположенные ближе к передним отделам мозга. Именно это наблюдали Эральдо Паулесу и его коллеги (см. рис. 2. 20)[271]. То же справедливо и для японского языка: различия в мозговой активности, вызванной словами на кандзи (требующей запоминания тысяч иероглифов) и кане (содержащей всего 46 слогов), могут быть объяснены заучиванием символов, размер и сложность которых разительно отличаются друг от друга[272].
Предпочтения нейронов и их роль в чтении
Хотя сегодня мы знаем, что процесс овладения чтением систематически задействует одну и ту же область коры, причина столь точной локализации до сих пор окутана тайной. Почему область «буквенной кассы» всегда расположена в левом полушарии и почему это верно для всех читателей и систем письменности? Что позволяет этой области обособиться от других зон, участвующих в распознавании объектов и лиц?
Хотя зрительная кора наделена значительной пластичностью, она не является «чистым листом», на котором отпечатываются результаты научения[273]. Сам факт того, что она организована в воспроизводимые субтерритории, расположение которых одинаково у всех людей, подразумевает, что она не эквипотентна. Что же формирует зрительный мозг читателя и объясняет стабильную локализацию процесса чтения? Очевидно, весь секрет в ранних предрасположенностях задействованных нейронов. По всей вероятности, будущим детекторам букв и биграмм изначально присущи особая зрительная чувствительность и проекции на речевые центры, делающие их особенно подходящими для овладения чтением.
|
|
|

Рис. 3. 10. Градиенты зрительной специализации объясняют, почему затылочно‑ височная область «буквенной кассы» находится в одном и том же месте у всех людей. Один такой градиент охватывает всю длину вентральной зрительной коры (вверху): латеральные области предпочитают изображения, представленные в центре сетчатки, или центральной ямке (показана темно‑ серым цветом), в то время как области ближе к срединной линии мозга выбирают изображения, отображаемые на периферии (показаны светло‑ серым цветом). Функции чтения и распознавания лиц, которые требуют тщательного анализа, реализуются корой, предпочитающей фовеальные зрительные стимулы (по материалам статьи Hasson et al., 2002). Использовано с разрешения Neuron.
Первый предрасполагающий фактор был выявлен Ури Хассоном, Рафи Малахом и их коллегами из Института Вейцмана[274]. Используя методы визуализации, они показали, что одни зрительные области преимущественно реагируют на изображения, представленные в центре сетчатки, а другие – на периферии. Соответствующий градиент предпочтений длиной в пять–шесть сантиметров пересекает всю вентральную зрительную кору (рис. 3. 10). Каждой ее точке, даже весьма удаленной от первичной зрительной области, свойственно выраженное предпочтение определенного места на сетчатке. Если области, расположенные по бокам мозга в обоих полушариях, предпочитают четкие визуальные стимулы, представленные близко к центральной ямке, то зоны, находящиеся ближе к срединной линии мозга, выбирают вход с периферии зрительного поля.
Происхождение этого градиента пока не установлено, но его размер и плавность напоминают те, что формируются в период эмбрионального развития. Возьмем, к примеру, постепенное уменьшение размера позвонков животного от спины к хвосту. Считается, что такой тип плавных изменений является результатом классического биологического механизма, который заключается в изменении концентрации химических сообщений – морфогенов, что распространяются по развивающемуся организму и определяют его базовый пространственный план[275]. Эта концепция восходит к математику Алану Тьюрингу. Тьюринг математически доказал, что если вещества вступают в химическую реакцию по мере их распространения через биологическую ткань, то они могут вызывать организованные в пространстве паттерны. Когда морфоген распространяется, он создает градиент убывающей концентрации, служащий суррогатным пространственным маркером. Так, если бы уровень морфогенов в центре мозга был выше, чем по бокам, и если бы эта концентрация модулировала притяжение нервных волокон, идущих из центральной ямки, результатом была бы генетически предвзятая нейронная сеть. Такая система в первую очередь обрабатывала бы стимулы с высоким разрешением, используя латеральные области коры. Именно такое пространственное устройство наблюдали Ури Хассон и его коллеги.
|
|
|
Но как этот градиент предпочтений сетчатки связан с чтением? Хассон заметил, что на нем область «буквенной кассы» всегда занимает одно и то же место. Оно характеризуется выраженным предпочтением четких фовеальных изображений. Соседняя область, преимущественно реагирующая на лица, тоже выбирает центральную ямку, тогда как зона, отвечающая за дома и пейзажи, предпочитает периферию зрительного поля.
Исходя из этих наблюдений, мы можем наметить простой сценарий, который объясняет, почему функция чтения реализуется именно в этом месте. С самого рождения зрительные нейроны уже неоднородны: одни, благодаря предпочтительным связям с центральной ямкой, предпочитают мелкие зрительные детали, другие – крупные стимулы. Естественно, первые лучше подходят для распознавания буквенных цепочек, требующих высокой четкости и остроты зрения. В результате функцию чтения преимущественно реализуют латеральные области зрительной коры, где такие нейроны наиболее многочисленны.
|
|
|
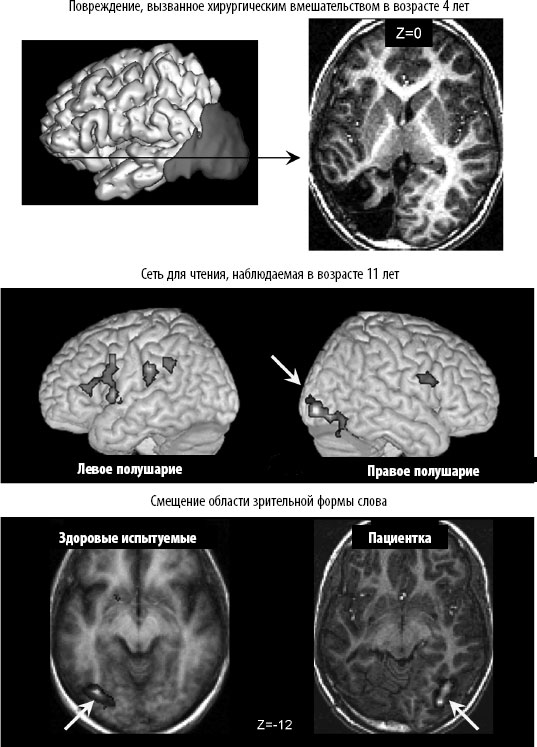
Рис. 3. 11. У детей сети для чтения гораздо более гибкие, чем у взрослых. У этой юной пациентки зона в левой затылочно‑ височной области, обычно связанная со зрительным распознаванием слов, была удалена хирургическим путем в возрасте четырех лет (травма хорошо видна на верхних снимках). У любого взрослого человека такое повреждение вызвало бы серьезное нарушение способности читать. Эта маленькая девочка научилась читать без особых сложностей. Семь лет спустя сеть чтения активировалась нормально, но зона зрительной формы слова поменяла полушария: теперь она находилась в правой затылочно‑ височной области, в месте, точно симметричном своему обычному расположению (внизу) (по материалам статьи Cohen et al., 2004). Использовано с разрешения Annals of Neurology.
Градиент предпочтений сетчатки, однако, представляет собой только одну из трех осей системы координат, определяющих местоположение области «буквенной кассы». Для выделения этой зоны необходимы и другие предпочтения. Как мы уже говорили, второй врожденный градиент идет от задних отделов мозга к передним и приблизительно очерчивает иерархический уровень каждой зрительной области (рис. 3. 5). Нейроны затылочного полюса реагируют на простые фрагменты изображений, в то время как передние участки коры предпочитают более сложные и структурированные объекты[276]. В этом отношении неудивительно, что область зрительной формы слова занимает относительно протяженную полосу коры, задняя часть которой реагирует на простые буквы, а передняя – на сложные фрагменты слов[277].
Наконец, важную роль могут играть различные свойства двух полушарий. Почему чтение главным образом опирается на левый зрительный мозг, а изображения лиц обычно передаются в правое полушарие? Отсутствие симметрии, или «нарушение симметрии», как говорят физики, может иметь зрительное происхождение. Хорошо известно, что левое полушарие преимущественно отвечает за различение малых локальных форм, а правое в основном обрабатывает глобальные формы[278]. Другим фактором может быть латерализация речи. Уже при рождении речь преимущественно обрабатывается в височной и лобной областях левого полушария. При формировании навыка чтения, вероятно, происходит отбор зрительных областей, обладающих наиболее многочисленными и прямыми проекциями на речевые центры. Если так, левая вентральная височная область будет иметь преимущество перед своим правополушарным двойником, аксоны которого могут попасть в левое полушарие только через мозолистое тело.
|
|
|
Иерархическая модель на рисунке 3. 9 предполагает, что смещение зрительного распознавания слов в левое полушарие – это последовательный процесс. Низкоуровневые области должны быть билатеральными: в обоих полушариях зрительные нейроны, отвечающие за первичную визуальную обработку, реагируют только на те буквы, которые представлены в противоположной половине зрительного поля. По мере продвижения вглубь мозга рецептивные поля расширяются, и мы находим клетки, способные реагировать на любую сторону зрительного поля. По моему убеждению, всякий раз, когда возникает такая потенциальная избыточность между полушариями, зрительные и лингвистические сдвиги заставляют левое полушарие брать верх над правым. Таким образом, чем выше уровень зрительной иерархии, тем больше нейронов в левом полушарии должны специализироваться на чтении.
Если мое предположение о врожденных предрасположенностях коры верно, то никакой заранее запрограммированной области для чтения не существует. Скорее, несколько генетически заданных сдвигов обеспечивают гамму нейронных предпочтений разных типов зрительных стимулов. Во время формирования навыка чтения зрительное распознавание слов просто выбирает участок коры, нейроны которого наиболее эффективно справляются с этой задачей. У всех людей пересечение генетически заданных градиентов создает оптимальное место для буквенных цепочек – область «буквенной кассы».
Что произойдет, если по той или иной причине эта область перестанет быть доступной? Благодаря научению функцию чтения должны взять на себя другие, менее эффективные, нейроны. Мой коллега Лоран Коэн и я недавно проверили эту гипотезу на одной юной пациентке с эпилепсией. В возрасте четырех лет девочка перенесла хирургическое удаление части мозга, в которой находилась часть зрительной системы, в том числе и участка, где обычно располагается область «буквенной кассы» (рис. 3. 11). Несмотря на столь серьезную операцию, она прекрасно научилась читать, хотя делала это немного медленнее, чем обычные дети. В возрасте 11 лет мы просканировали ее мозг и наконец поняли, как ей это удалось. Несмотря на то, что все области левого полушария, отвечающие за устную речь, остались нетронутыми, она компенсировала потерю «буквенной кассы», распознавая письменные слова с помощью интактного правого полушария. Более того, чтение избирательно активировало зону, в точности симметричную области «буквенной кассы» у здоровых людей: в латеральной затылочно‑ височной борозде, в том же самом месте как по латеральной, так и по передне‑ задней оси, только в правом полушарии (рис. 3. 1)[279]. Иначе говоря, ее «буквенная касса» просто «переехала» из левого полушария в правое!
Другие исследователи наблюдали аналогичную активацию правого полушария у ребенка с гиперлексией, который очень рано научился читать[280]. Правостороннее расположение области зрительной формы слова также встречается у взрослых пациентов с чистой алексией, которые перенесли инсульт, затронувший левую зрительную систему[281]. Таким образом, вклад этой области в чтение далеко не исключительный. Складывается впечатление, что у зрительной системы, в силу изначально присущих ей предрасположенностей, имеется своего рода список областей, наиболее подходящих для чтения. Если первый кандидат больше не доступен, задействуется кандидат номер 2 – симметричная область в правом полушарии.
Никакое другое экспериментальное открытие не могло бы лучше проиллюстрировать фундаментальный вывод о том, что заранее заданной области для чтения не существует. Овладение чтением предполагает минимальную перестройку нашего мозга приматов. Используя все доступные средства, он приспосабливает наиболее подходящие области зрительной коры для решения новой задачи распознавания слов.
|
|
|
