 |


Биоэнергетика катаболизма жирных кислот
|
|
|
|
Биоэнергетика катаболизма жирных кислот
с нечетным числом углеродных атомов
Первоначальное уравнение для расчета биоэнергетики окисления жирных кислот с нечетным числом углеродных атомов имеет вид
n = (m – 3)/2∙ 10 + (m – 3)/2∙ 4 + 25 – 2 – 1. (6)
n – количество запасаемых молекул АТФ.
m – количество углеродных атомов жирной кислоты.
(m – 3)/2 – количество ацетил-КоА образующихся в процессе β -окисления.
10 – количество эквивалентов АТФ образующихся в цикле Кребса при окислении 1 молекулы ацетил-КоА.
(m – 3)/2 – число этапов β -окисления.
4 – количество эквивалентов АТФ образующихся за 1 этап (1НАДН + Н+, 1 ФАДН2).
25 – количество эквивалентов АТФ образующихся при полном окислении сукцинил-КоА (промежуточный метаболит) за 2, 5 оборота цикла Кребса.
2 – количество АТФ потраченных на активирование жирной кислоты.
1 – количество эквивалентов АТФ потраченных в карбоксилазной реакции.
Упрощенное уравнение 6 принимает вид: n = 7m + 1. (7)
Уравнение 7 справедливо для подсчета биоэнергетики полного окисления жирных кислот, с нечетным числом углеродных атомов начиная с m = 3.

Рис. 14. Катаболизм гептановой кислоты
Катаболизм разветвленных жирных кислот
Катаболизм жирных кислот изостроения с разветвлением на ω -конце молекулы осуществляется с использованием ферментов участвующих в их активировании, транспорте в митохондрии, β – окисления, цикла Кребса, окислительного фосфорилирования. Однако в процессе их окисления используются и дополнительные ферментативные реакции. Эти реакции необходимы для превращения изобутирил-КоА (образуется в процессе β – окисления разветвленных жирных кислот с четным числом углеродных атомов) и изовалерил-КоА (образуется в процессе β – окисления разветвленных жирных кислот с нечетным числом углеродных атомов) до промежуточных продуктов цикла Кребса. Изобутирил-КоА превращается в сукцинил-КоА, а изовалерил-КоА – в три молекулы ацетил-КоА. Окисление изовалерил-КоА представлено на рисунке 15.
|
|
|
Энергетический баланс окисления
разветвленных жирных кислот
Для расчета энергетического баланса жирной кислоты изостроения с нечетным числом углеродных атомов можно воспользоваться выражением
n = (m – 5)/2∙ 10 + (m – 5)/2∙ 4 + 30 + 1, 5 – 1 – 2.
После преобразования этого выражения получим уравнение
n = 7m – 6, 5.
Полученное уравнение можно использовать для расчета биоэнергетики окисления жирных кислот изостроения с нечетным числом углеродных атомов.
Для расчета биоэнергетики окисления жирных кислот с четным числом углеродных атомов первоначальное выражение имеет вид
n = (m – 4)/2∙ 10 + (m – 4)/2∙ 4 + 31, 5 – 2.
После преобразования получаем
n = 7m + 0, 5.

Рис. 15. Катаболизм изовалерил-КоА до ацетил-КоА
Метаболизм глицерола

Примечание: ОкФ – окислительное фосфорилирование, ЦПЭ – цепь переноса электронов.
Рис. 16. Катаболизм глицерола
Глицерол пищевых и эндогенных глицеролипидов может участвовать не только для получения энергии, но и для других целей, как это показано на рисунке 17.

Рис 17. Метаболизм глицерин
Анаболизм липидов
Синтез жирных кислот
Синтез жирных кислот осуществляется на мембранах гладкого эндоплазматического ретикулума (ГЭР) ассоциированным с ним мультиферментным коньюгатом синтетазы жирных кислот. Строительным материалом для синтеза жирных кислот является ацетил-КоА. Это соединение образуется в митохондриях в процессе окисления углеводов, липидов, аминокислот, однако в синтезе, главным образом, участвует ацетил-КоА, образованный из углеводов. Внутренняя мембрана митохондрий непроницаема для ацетил-КоА, поэтому для его выведения в цитозоль используется цитратный челнок, показанный на рисунке 18.
|
|
|
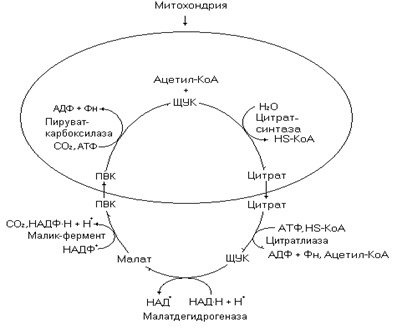
Рис. 18. Цитратный челнок
В работе челночного переноса участвует фермент цикла Кребса - цитратсинтаза, катализирующий реакцию конденсации ацетил-КоА и щавелевоуксусной кислоты (ЩУК) с образованием цитрата. Полученное соединение транспортируется из митохондрий транслоказой, локализованной во внутренней мембране митохондрий. В цитозоле цитрат распадается до ЩУК и ацетил-КоА в цитратлиазной реакции. В реакции участвуют также АТФ и НS-КоА. Таким образом, митохондриальный ацетил-КоА оказывается в цитозоле, где используется в синтезе жирной кислоты. ЩУК образованный в этой же реакции, преобразуется в малат цитоплазматической малатдегидрогеназой. В свою очередь малат декарбоксилируется до пировиноградной кислоты (ПВК) малик-ферметом (малатдегидрогеназа декарбоксилирующая). ПВК транспортируется в цитозоль, где кабоксилируется до ЩУК. На этой реакции цикл завершается, следовательно, ЩУК выполняет функцию молекулы переносчика ацетил-КоА, не расходуясь при этом.
Мультиферментный коньюгат синтазы жирных кислот человека участвует в синтезе пальмитиновой кислоты – С16, состоящий из двух димеров. Одна полипептидная цепь коньюгата содержит семь каталитических центров и ацилпереносящий белок (АПБ).
В синтезе пальмитата участвует малонил-КоА, образующийся путем карбоксилирования ацетил-КоА.
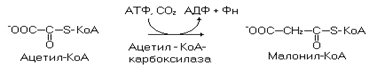
Синтез пальмитата идет поэтапно (семь этапов), в повторяющейся последовательности реакций (рис. 19).

Рис. 19. Этап синтеза пальмитиновой кислоты и переход к новому этапу.
Каждый этап состоит из шести реакций. В первой реакции идет перенос ацетильного остатка на ацил-переносящий белок (АПБ). Образуется ацетил-АПБ, в котором ацетильный остаток связан с серой остатка цистеина. Далее образовавшийся комплекс взаимодействует с малонил-КоА, который связывается с серой остатка фосфопантетеина АПБ. В третьей реакции идет перенос ацетильного остатка на малонильный, который при этом декарбоксилируется. Углекислый газ, образованный в этой реакции тот же самый, который участвовал в синтезе малонил-КоА. Три последние реакции этапа включают две реакции гидрирования с участием НАДФН + Н+ и одну реакцию дегидратации. Восстановленная форма кофермента (НАДФН + Н+) поступает из пентозофосфатного пути или реакции катализируемой малик-ферментом. Этап завершается образованием остатка масляной кислоты связанной с АПБ (бутирил-S-АПБ). Остаток бутирата переносится на серу цистеина и начинается новый этап, но уже без участия ацетил-КоА.
|
|
|
На каждом этапе синтеза жирной кислоты имеют место две реакции восстановительного синтеза, донором водорода в которых используется восстановленная форма пиридинового кофермента – НАДФН. Основными источниками НАДФН являются пентозофосфатный путь, реакции катализируемые малик-ферментом и изоцитратдегидрогеназой цитоплазматической.
|
|
|
