 |


Восприятие (ощущение) боли включает в себя физиологические процессы: трансдукцию, трансмиссию, модуляцию и перцепцию.
|
|
|
|
Восприятие (ощущение) боли включает в себя физиологические процессы: трансдукцию, трансмиссию, модуляцию и перцепцию.
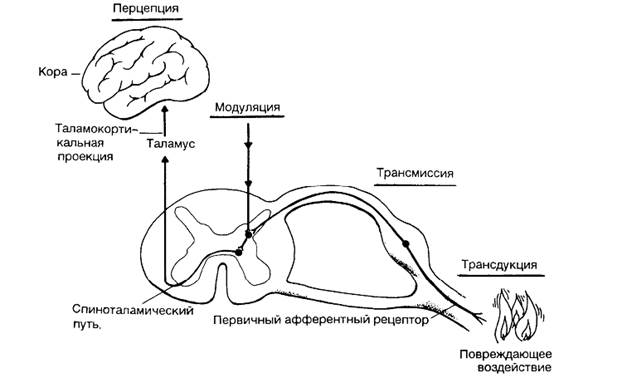
Трансдукция – процесс, при котором повреждающее воздействие трансформируется в виде электрической активности на окончаниях чувствительных нервов.
Процесс трансдукции может инициироваться и усиливаться в результате нескольких механизмов:
- прямая активация ноцицепторов;
- сенсибилизация ноцицепторов с последующим усилением их активности;
- выход аллогенных веществ из плазмы.
Трансдукция в глубоких структурах (мышцы, суставы) по своим механизмам аналогична трансдукции в коже. Проведение боли обеспечивают два основных типа волокон. Присутствие двух путей проведения объясняет наличие и двух основных компонентов боли: «первичной боли», развивающейся всегда быстро и остро, имеющей всегда четкую локализацию и проводимую А-δ -волокнами, а также «вторичной» боли, которая проводится С-волокнами и имеет всегда медленное начало и нечеткую локализацию. Миелинизированные А-δ -волокна имеют диаметр 2–5 мкм и передают импульсы со скоростью от 12 до 30 м/сек, тогда как немиелинизированные C-волокна имеют диаметр 0, 4–1, 2 мкм, а скорость проведения импульсов по ним равна 0, 5–2 м/сек. И те и другие волокна оканчиваются в задних рогах спинного мозга. Иннервация костной ткани осуществляется волокнами Аδ и С, образующих сплетения в надкостнице. В корковом слое и костном мозге ноцицептивных волокон нет. Костная ткань обладает самым низким болевым порогом из всех глубоких соматических структур. Наблюдения во время операций показали, что внутренние органы относительно менее чувствительны к стимуляции (разрез, ожог, сжатие), чем наружные покровы. Однако, эти органы восприимчивы к смещениям типа скручивания и растяжения. Их иннервация обеспечивается волокнами типа А δ и С. Висцеральные афференты обычно проходят вместе с симпатическими волокнами и снабжены большими сливающимися полями рецепции.
|
|
|
Передача импульсов по периферической нервной системе называется трансмиссией. Трансмиссия – проведение возникших импульсов по системе чувствительных нервов.
Проводящие пути, ее обеспечивающие, сформированы из трех компонентов:
- первичного афферентного нейрона, расположенного в ганглиях заднего корешка, доходящего до спинного мозга;
- восходящего промежуточного нейрона, простирающегося от спинного мозга до ствола головного мозга и таламуса;
- таламокортикальных проекций.
Клеточные тела афферентных нейронов расположены в ганглиях заднего корешка. Периферический отросток этих клеток выходит в составе заднего корешка, а центральный идет к спинному мозгу в проксимальном отрезке этого же корешка. Задний корешок, подходя к спинному мозгу, расщепляется на 12-15 пучков. Немиелинизированные волокна при этом смещаются в передне-латеральном направлении, большие миелинизированные – в заднемедиальном направлении. Большие волокна проникают в спинной мозг медиально и сразу раздваиваются на восходящую и нисходящую ветви в составе заднего столба. Малые волокна входят в спинной мозг латерально и также разделяются на восходящую и нисходящую ветви, которые распространяются на 1-2 сегмента от места вхождения. Большинство этих волокон А-δ и С проходят в составе тракта Лиссауэра, который в виде ленты накрывает задний рог. Серое вещество спинного мозга по своей структуре представляет слои или пластины. Они пронумерованы римскими цифрами в порядке их расположения, начиная от дорсальной поверхности. Весь задний рог сформирован пластинами I-VI. Передний рог сформирован пластинами VII-X. Волокна С заканчиваются преимущественно в пластинах I и II. Окончания ноцицепторов А-δ находятся в пластинах I и V и, частично, в пластине Х. Афферентные волокна внутренних органов оканчиваются преимущественно в пластинах I и V. Пластины I-II-V играют ведущую роль в ноцицептивной трансмиссии. Большинство афферентных волокон заканчиваются в заднем роге с той же стороны, но некоторые из них пересекают серое вещество сразу же позади центрального канала и заканчиваются в заднем роге противоположной стороны. Задние рога спинного мозга называют еще и «воротами боли», где ноцицептивные импульсы способны меняться (или переключаться). Концепция о заднем роге как о воротах, которые могут быть «закрыты» с помощью фармакологического воздействия на процессы трансдукции, трансмиссии и модуляции, формируют основополагающий физиологический принцип эффективного преодоления интраоперационной и послеоперационной боли. В нейротрансмиссии на уровне заднего рога важное значение имеет частота поступающих импульсов с периферии. При частоте ноцицептивных сигналов более 3 в одну секунду постсинаптический разряд становится более продолжительным, что способствует усилению болевого ощущения. Этот феномен временной суммации получил название «windup» (взвинчивание).
|
|
|
Вторые нейроны функционально представляют собой либо ноцицептивно-специфические нейроны, либо нейроны с широким динамическим диапазоном. Ноцицептивно-специфические нейроны реагируют исключительно на ноцицептивные стимулы. В норме эти нейроны находятся в состоянии покоя и реагируют только на высокопороговую ноцицептивную стимуляцию. Нейроны же широкого диапазона воспринимают не только ноцицептивные, но и неноцицептивные афферентные импульсы. Нейроны широкого диапазона реагируют на менее интенсивную болевую стимуляцию.
От второго нейрона возбуждение может пойти двумя путями:
1. Специфический (лемнисковый). Аксоны вторых нейронов в составе спиноталамического тракта идут к специфическим ядрам таламуса. В таламусе возбуждение передается на третий нейрон. Аксон третьего нейрона достигает коры головного мозга. Особенность специфических ядер таламуса состоит в том, что они передают возбуждение «прямо по назначению» в нужную зону коры.
|
|
|
2. Неспецифический (экстралемнисковый). Начинается от второго нейрона спинного мозга и по коллатералям идет в различные структуры мозга. В зависимости от места окончания выделяют три основных тракта – неоспиноталамический (спинной мозг-таламус), спиноретикулярный, спиномезенцефалический (спинной мозг – средний мозг). Возбуждение по этим путям поступает в неспецифические ядра таламуса и оттуда во все отделы коры больших полушарий. Особенность неспецифических ядер таламуса как раз и заключается в том, что они обеспечивают обширные связи таламуса с разными структурами головного мозга.
Аксоны большинства вторых нейронов, направляясь контралатерально, пересекают среднюю линию спинного мозга в передней белой спайке и, устремляясь рострально, образуют спиноталамический тракт. Он является важнейшим восходящим путем, существующим для передачи широкого спектра информации о свойствах болевого стимула.
Спиноталамический (неоспиноталамический) тракт оканчивается в таламусе, ретикулярной формации, ядре шва и околоводопроводном сером веществе. Спиноталамический тракт состоит из латеральной и медиальной частей. Латеральный спиноталамический тракт идет главным образом к вентральным заднелатеральным ядрам таламуса и отвечает за локализацию, интенсивность и продолжительность боли.
Медиальный спиноталамический (палеоспиноталамический) тракт, состоящий из спиноретикулярного и спиномезенцефалического трактов, несет импульсы к среднему таламусу. Он обеспечивает восприятие вегетативных и эмоциональных аспектов боли.
Значительный объем ноцицептивной информации поступает в ствол мозга через аксоны спиноретикулярного тракта, являющегося вторым по величине путем передачи ноцицептивной информации. Терминали этих аксонов распределены в медиальной ретикулярной формации продолговатого мозга, а также в релейных ядрах таламуса. Некоторые спиноретикулярные нейроны являются энкефалинсодержащими. Через систему спиномезенцефального тракта запускаются сложные соматические и висцеральные антиноцицептивные рефлексы.
|
|
|
Терминали нейронов спиноталамического тракта оканчиваются в специфических (релейных) ядрах таламуса. Современные морфологические, физиологические исследования и обширная практика нейрохирургических вмешательств свидетельствуют о том, что ноцицептивная информация достигает высших отделов мозга через многочисленные дублирующие каналы, которые в силу обширной конвергенции и диффузных проекций вовлекают в формирование боли сложноорганизованную иерархию различных структур головного мозга, в которых происходит взаимодействие разномодальных афферентных систем. Ноцицептивный поток информации передается из спинного мозга практически во все структуры головного мозга: ядра ретикулярной формации, центральное околоводопроводное серое вещество, таламус, гипоталамус, лимбические образования и кору больших полушарий, которые выполняют самые разнообразные функции как сенсорного, двигательного, так и вегетативного обеспечения защитных реакций, возникающих в ответ на ноцицептивное раздражение. Так, ретикулярная формация, очевидно, принимает участие в формировании афферентно-мотивационного компонента боли и в интеграции боли с автономными и двигательными функциями. Гипоталамус в качестве регулятора автономной нервной системы определяет автономные и эндокринные реакции в ответ на все виды стимуляции, включая боль. Нейроны, содержащие вазопрессин (антидиуретический гормон – АДГ) и кортикотропин-рилизинг-гормон, локализуются в паравентрикулярном ядре гипоталамуса. Лимбическая система обеспечивает аффективно-мотивационный аспект перцепции боли.
Модуляция – это процесс, при котором ноцицептивная трансмиссия модифицируется под влиянием нейрональных воздействий. Модуляция может как ослабить, так и усилить боль.
Модуляция боли происходит как на периферии в ноцицепторах, в спинном мозге, так и в центральных супраспинальных структурах.
Периферическая модуляция. При повторном возбуждении чувствительность ноцицепторов и их нейронов возрастает. Этот феноменполучил название сенситизации. Повышение чувствительности ноцицепторов приводит к снижению порога возбуждения, увеличению частоты импульсации при стимуле той же интенсивности, уменьшению латентного периода, или времени ожидания, и появлению спонтанной активации после прекращения стимуляции. Данное явление, именуемое первичной гипералгезией (повышение чувствительности к боли), опосредуется такими альгогенами, как гистамин, брадикинин, простагландин-Е2, а также лейкотриенами, высвобождаемыми при повреждении тканей.
|
|
|
Вторичная гиперальгезия, или нейрогенное воспаление, проявляется соответствующей триадой: гиперемией, местным отеком и повышенной чувствительностью к ноцицептивным стимулам. Вторичная гиперальгезия обусловлена главным образом высвобождением субстанции-P из коллатеральных аксонов первых афферентных нейронов. Субстанция-P вызывает высвобождение гистамина и серотонина, локальную вазодилатацию, отек тканей и выработку лейкотриенов.
Центральная модуляция.
Модуляция может, как усиливать, так и блокировать боль. К механизмам, усиливающим боль, относят:
1. Феномен ≪ взвинчивания≫, или ≪ реакция испуга≫, и сенситизация вторых нейронов.
2. Расширение рецепторных полей.
3. Гипервозбудимость сгибательных рефлексов.
К нейромедиаторам центральной сенситизации относятся субстанция-P, пептид, ассоциированный с геном кальцитонина (ПАГК), вазоактивный интестинальный пептид (ВИП), холецистокинин, ангиотензин, аланин, L-глутамат и L-аспартат. Данные вещества изменяют возбудимость мембраны нейрона за счет взаимодействия с рецепторами, связанными с G-белком, и активации внутриклеточных вторичных мессенджеров, фосфорилирования белков и увеличения внутриклеточной концентрации кальция. Глутамат и аспартат активируют N-метил-D-аспартат-рецепторы (NMDA). Стимуляция NMDA-рецепторов вызывает повышение в нейронах внутриклеточного Ca2+, что активирует NO-синтетазу и продукцию оксида азота (NO).
Оксид азота принимает непосредственное участие в развитии гипервозбудимости, проявляющейся гиперальгезией или аллодинией (болезненный ответ на обычно неболезненный стимул), увеличивая уровень нейротрансмиттеров в центральных терминалях.
Перцепция (от лат. Рercipere -восприятие) Третьи нейроны расположены в таламусе и посылают волокна до соматосенсорных полей I и II в постцентральной извилине и верхней стенке сильвиевой борозды.
В данных областях коры обеспечиваются перцепция и дискретная локализация боли. Р. Мелзак (1981) в разделе о психологии боли указывает, что боль относится к категории сложных переживаний, а не является простым ощущением, вызванным специфическим раздражителем.
|
Структура низкомолекулярных нейротрансмиттеров | ||
| Характер действия | Основная функция | |
| возбуждение | ингибирование | |
| нейромедиаторы | 
| 
|
| нейромодуляторы | 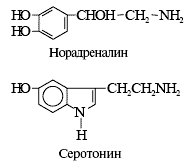
| 
|
Научные данные подтверждают взгляд на боль как на перцептуальное явление, на которое влияет прошлое каждого человека, оценка обстоятельств, вызывающих боль, и психическое состояние в данный момент.
Таким образом, боль становится функцией человека в целом, включая и его мысли, и страхи в данный момент, и его надежды на будущее.
|
|
|
