 |


Природа мембранных потенциалов
|
|
|
|
ПРИРОДА МЕМБРАННЫХ ПОТЕНЦИАЛОВ
Потенциал покоя
Все живые клетки имеют мембранный потенциал (МП), который определяется разностью зарядов между наружной и внутренней поверхностями мембраны и измеряется в милливольтах (мВ). МП возникает в результате разделения структурами мембраны заряженных ионов. Постоянная работа Na-K АТФазы по переносу ионов через мембрану приводит к тому, что в состоянии покоя концентрация калия в 48 раз превышает наружную, а снаружи концентрация натрия примерно в 10 раз выше внутренней. В мембране существуют так называемые калиевые каналы «утечки», благодаря которым проницаемость мембраны в состоянии покоя к ионам К+ много больше, чем ко всем остальным ионам. В условиях когда внутри клетки концентрация К+ много больше, чем снаружи, а мембрана обладает избирательной проницаемостью к калию, эти катионы должны по концентрационному градиенту выходить из клетки. Выход каждого иона К+ сопровождается выносом из клетки одного положительного заряда. При этом анионы, не способные покинуть клетку, способствуют возникновению отрицательного заряда на внутренней поверхности мембраны. Это ведет к созданию электрического градиента, направленного для положительных зарядов внутрь клетки. Выход ионов К+ из клетки возможен только до того момента, когда энергия концентрационного градиента, выталкивающая калий из клетки, не будет уравновешена энергией электрического градиента, препятствующей этому процессу. Потенциал, возникающий при этом на мембране, называется равновесным калиевым потенциалом и описывается уравнением Нернста.
EK = RT ln [K]in
ZF [K]out
|
|
|
где R – универсальная газовая постоянная;
Т - абсолютная температура (Ко );
Z - валентность иона;
F – константа Фарадея
[K]in , [K]out – внутри и внеклеточная концентрации ионов калия.
Для каждого из ионов, присутствующих в физиологически значимых концентрациях в внутри и внеклеточных средах, могут быть вычислены соответствующие равновесные потенциалы.
Если подставить в уравнение Нернста реальные концентрации ионов калия, то расчетное значение равновесного потенциала будет -95 мВ, однако при измерении потенциала в клетке при помощи микроэлектрода величина его не превышает -90 мВ. Объясняется это тем, что мембрана не идеальна и в состоянии покоя имеет небольшую проницаемость и для других ионов, за счет чего снижается абсолютное значение мембранного потенциала.
В покое проницаемости к таким ионам описываются следующими соотношениями.
PK: PNa: PCl = 1. 0: 0. 04: 0. 45
Потенциал покоя (ПП) – это потенциал, устанавливающийся в клетках в состоянии покоя и являющийся результатом работы Na-K насоса по разделению ионов и зарядов относительно мембраны и результатом высокой избирательной проницаемости мембраны к ионам К+.
Дляизмерения мембранных потенциалов используют микроэлектроды, представляющие собой стеклянные микропипетки с диаметром кончика менее 0, 5 микрометра. Столь малый диаметр кончика гарантирует от повреждения мембрану при ее проколе микроэлектродом. Микроэлектрод заполнен раствором электролита (3 М раствор KCl) для снижения электрического сопротивления. Микроэлектрод посредством тонкой подвижной платиновой проволоки соединен с электронным усилителем и устройством отображения и регистрации потенциалов (компьютером) (Рис. 2).
В покое клетка находится в состоянии поляризации (ПП= -90мВ). Уменьшение потенциала покоя по абсолютной величине называется деполяризацией, увеличение – гиперполяризацией.
|
|
|

Рис. 2. Регистрация внутриклеточного потенциала при помощи микроэлектрода.
1- микроэлектрод, введенный в клетку и соединенный с усилителем,
2- измерение мембранного потенциала на экране осциллографа.
Потенциал действия
Потенциал действия (ПД) – кратковременное изменение мембранного потенциала, связанное с открытием ионных каналов, способное к самостоятельному распространению по возбудимым клеткам и являющееся сигналом, посредством которого передается информация.
В мембране помимо насосов и каналов утечки существуют другие ион-селективные каналы, участвующие в процессе возбуждения. Наибольшее значение для возникновения и проведения ПД имеют натриевые и калиевые каналы. Каналы представляют собой трансмембранные интегральные белки, состоящие из нескольких тысяч аминокислот, формирующие ионную пору, пронизывающую всю толщину мембраны. Каждый канал содержит ион селективный фильтр, пропускающий только определенные ионы, и систему воротных механизмов, обеспечивающих активацию или инактивацию канала (Рис. 3).

Рис. 3 Последовательность работы воротных механизмов натриевых и калиевых каналов при возбуждении мембраны.
Натриевый канал управляется двумя воротными механизмами. На внешней стороне мембраны находится «вольт-зависимый» (или электро-зависимый) механизм (m-ворота), который открывается при определенной степени деполяризации мембраны. На внутренней стороне мембраны расположен «время-зависимый» механизм (h-ворота), который закрывает канал через определенное время (как правило 1 мс) после полного открытия m-ворот и, наоборот, открывается после полного закрытия «вольт-зависимых» ворот.
Таким образом, натриевый канал может находиться в трех состояниях:
1. Закрытое состояние – состояние покоя, когда m-ворота закрыты, h-ворота открыты. В этом состоянии при деполяризации канал может быть открыт.
2. Активированное (открытое) состояние (оба воротных механизма открыты).
3. Инактивированное состояние, при котором m-ворота открыты, а h-ворота закрыты. В этом состоянии канал не может быть активирован и открыт, пока не перейдет в закрытое состояние.
|
|
|
Калиевый канал имеет один воротный механизм – так называемые n-ворота, которые открываются при полной деполяризации мембраны. Закрытие ворот происходит при реполяризации до ПП то есть до значений потенциала на мембране от -70 до -90 мВ. Канал не имеет состояния инактивации и может быть только в открытом или закрытом состоянии.
В покое все описанные выше электрозависимые каналы закрыты, но благодаря калиевым каналам утечки проницаемость мембраны для ионов К+ высока и ее МП близок к значениям равновесного калиевого потенциала (ЕК). При деполяризации мембраны (за счет электротонического проведения тока от соседних возбужденных клеток или в результате приложения тока от электростимулятора) происходит сдвиг МП до значений меньших по абсолютной величине, чем -60 мВ. Это вызывает, пропорциональное сдвигу потенциала открытие активационных (m) ворот и поступлению в клетку положительно заряженных ионов Na+. Таким образом, возникает регенеративный процесс с положительной обратной связью, когда чем больше деполяризация под действием Na+, тем больше открываются ворота и больше поток входящего натрия. В этот момент ионы Na+ входят в клетку под действием однонаправленных сил концентрационного и электрического градиентов. Ток, текущий по Na+ каналам называется «быстрым» натриевым током и определяет фазу быстрой деполяризации потенциала действия, длительность которой не превышает 1 мс. Мембранный потенциал, при этом, стремится к значениям равновесного натриевого потенциала (+48 мВ), но не достигает его, поскольку примерно через 1мс после открытия m ворот происходит закрытие h-ворот и натриевые каналы оказываются инактивированными. Процесс этот происходит при значении МП около +30 мВ. Превышение потенциала над нулевым уровнем называется «овершут».
В этот момент открываются калиевые каналы. На этот раз ионы калия так же находятся под действием выталкивающих сил не только концентрационного, но и электрического градиентов. Выходящий калиевый ток определяет фазу реполяризации ПД (Рис. 4). Ток калия из клетки настолько интенсивен, что по окончании ПД, мембранный потенциал еще некоторое время возрастает по абсолютной величине. Это увеличение называется фазой следовой гиперполяризации. Затем происходит закрытие калиевых каналов и МП возвращается к значению потенциала покоя. В конце фазы реполяризации происходит переход натриевых каналов из состояния инактивации в закрытое состояние. При этом m-ворота закрываются, а h- ворота открываются. С этого момента канал может быть снова активирован при деполяризации. Все время в течение которого Na-каналы инактивированы генерация ПД в клетке невозможна и клетка находится в состоянии абсолютной рефрактерности (невозбудимости). В фазу следовой гиперполяризации клетка может быть возбуждена, но только сверхпороговым импульсом, превышающим по своей амплитуде обычный порог возбуждения. Это состояние называется относительной рефрактерностью.
|
|
|
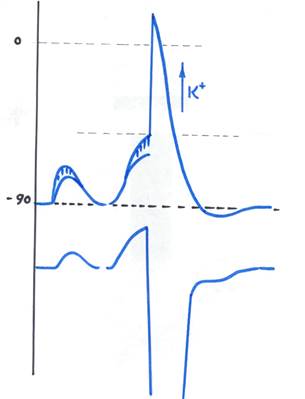
Рис. 4 Потенциал действия и локальный ответ. Изменение возбудимости в ходе ПД.
Заштрихованные области отражают вклад входа Na через каналы при допороговой деполяризации. Ниже приведен график изменения возбудимости в ходе локального ответа и ПД
Уровень деполяризации клетки при котором происходит необратимое открытие натриевых каналов называется уровнем критической деполяризации. Разность между ПП и уровнем критической деполяризации будет порогом возбуждения. На рис. 4 приведен график изменения возбудимости клетки в различные фазы ПД.
При деполяризации клетки ниже порога на мембране также возникает потенциал, но в отличие от ПД он не способен достичь порога возбуждения в соседних областях клетки или соседних клетках. Такой потенциал распространяется на короткое расстояние и быстро затухает. Поэтому он называется локальным ответом.
Потенциал действия подчиняется так называемому закону «все или ничего». Под понятием «все» понимают сам ПД, под понятием «ничего» - локальный ответ. Поэтому любая деполяризация мембраны, превышающая уровень критической деполяризации вызывает генерацию ПД.
Процесс распространения возбуждения по нервному волокну достаточно медленный. Скорость проведения значительно возрастает, если волокно миэлинизировано. Шванновские глиальные клетки не только защищают нервное волокно от механических повреждений, но за счет своего высокого электрического сопротивления обеспечивают быстрое проведение деполяризующего тока по внутреннему просвету волокна (Рис. 5). При этом, ПД как бы перепрыгивают с одного перехвата Ранвье на другой. Генерация ПД в каждом перехвате Ранвье обеспечивает сверхпороговую деполяризацию в следующем перехвате и быстрое движение нервного импульса вдоль волокна. В табл. 1 приведены такие скорости для разных типов волокон.
|
|
|

Рис. 5 Проведение локального ответа (вверху) и потенциала действия по обычному (немиэлинизированному) (средний рисунок) и миэлинизированному (внизу) волокну.
|
|
|
