 |


Карогидные тельца. Типы клеток в каротидном тельце. Функция каротидных телец
|
|
|
|
Карогидные тельца
В то время как мониторинг РС()2 происходит в стволе головного мозга, отслеживание Р( )2 является функцией исключительно каротидныхтелец (рис. 16-3). Они расположены у бифуркации общих сонных артерий на внутреннюю и наружную (рис. 16-3 А). Несмотря на свой миниатюрный размер, каротидные тельца получают огромный кровоток - от 1. 4 до 2. 0 л/мин на 100 г ткани, что соответствует особой роли этого органа, как главного датчика, чувствительного к колебаниям кислорода в артериальной крови.
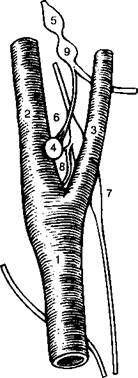

Рис. 16-3. Каротидиос тельце. (А) бифуркация правой сонной артерии (вид спереди). Общая сонная артерия (1) делится па наружную (2) и внутреннюю (. 4). Каротидное тельце (4) иппсрвирустся чувствительными волокнами, исходящими из каменистого ганглия (5) и проходящими в составе нерва каротидпого синуса (ft). Верхний шейный ганглий (7) тоже интернирует каротидное тельце посредством гапглиогломсрулярных нервов (8). Узловатый ганглий (9) расположен снаружи внутренней сонной артерии. (Б) Каротидиое тельце, состоит из хеморецепторпых (1) и поддерживающих (2) клеток. В цитоплазме хсморецепторпых клеток находятся сииаптические пузырьки (Л), которые тесно соприкасаются с чувствительными окончаниями (4) ветвящихся волокон нерва каротидпого синуса (5). Вся совокупность этих клеток окружена капиллярами (ft). (Из: (ion/ale/. С'., Alinarax L, Obeso Д., Ritual R. Oxygen and acid chemorcccption in the carotid body chemoreceptors. Trends in Neuroscicncc 15: I/I ft-153/1992. )
Типы клеток в каротидном тельце
Каротидные тельца состоят из клеток нескольких типов (рис. 16-ЗБ). Главной из них является гломусная клетка (I тип). Она содержит несколько нейротрансмит-теров, включая большое количество допамина и других катехоламинов, а так же серотонин, ацетилхолин и некоторые нейропептиды. Гломусные клетки являются настоящими хемочувствительными клетками.
|
|
|
Группы гломусных клеток окружены поддерживающими клетками (II тип). На одну поддерживающую клетку приходится от трех до шести гломусных. Физиологическая роль поддерживающих клеток неизвестна. Считается, что эти клетки ультраструктурно и функционально связаны с глией центральной нервной системы. Группы гломусных клеток и окружающие их поддерживающие клетки снабжены богатой сетью капилляров, которые обеспечивают оптимальные условия для их хе-мосенсорной функции относительно кислорода.
Функция каротидных телец
v
Гломусные клетки образуют синапсы с афферентными волокнами нерва каро-тидного синуса. Чувствительная ветвь языкоглоточного нерва и нерв каротидного синуса иннервируют каротидное тельце и барорецепторы каротидного синуса. Афферентные волокна языкоглоточного нерва берут свое начало в чувствительных нейронах каменистого ганглия. В ответ на артериальную гипоксемию гломусные клетки высвобождают допамин пропорционально степени снижения РаО2. Высвобождение допамина модулируется отклонениями рН, вызванными метаболическим или дыхательным ацидозом, и приводит к значительному повышению тонической активности афферентных чувствительных волокон каротидных телец. Эти афферентные волокна проявляют некоторую степень тонической активности в условиях нормок-сии и даже гипероксии. Они " замолкают" только в присутствии выраженной артериальной гипероксии и гипокапнии (рис. 16-4).
Когда РаО2 падает ниже 60 мм рт. ст., активность афферентных нервов каротидного тельца возрастает экспоненциально (рис. 16-4 А). Поскольку частота импульса-ции единичного афферентного волокна каротидного тельца составляет от 5 до 25 импульсов в секунду, каротидное тельце быстро отвечает на изменения РаО2, происходящие в течение одного дыхательного цикла. ,
|
|
|
Важнейшее значение каротидного тельца в регуляции оксигенации подтверждается тем, что после его двустороннего удаления или денервации гипоксический вентиляторный ответ у человека полностью исчезает. У некоторых других биологических видов подобные кислород-чувствительные органы присутствуют также в дуге аорты. Этим аортальным тельцам свойственна реактивность, функционально подобная реактивности каротидных телец.
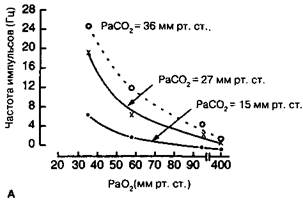
Рис. 16-4. Активность каротидного тельца как функция Рао2 или Расо2. (А) Активность одиночного афферентного волокна каротидного тельца как функция Paov при трех различных уровнях Расо2. (Б) Активность одиночного афферентного волокна каротидного тельца как функция Расо, при четырех различных уровнях Рао2. (Из: Cherniack N. S, Pack A. I. Control of ventilation. In: Fishman A. P., ed. Pulmonary Diseases and Disorders. New York: McGraw-Hill, 1988: 133. )

В отличие от решающей роли каротидных телец в регуляции оксигенации, их участие в формировании вентиляторного ответа на изменение РаС()2 сравнительно невелико. Несмотря на то, что частота импульсации афферентных нервов каротид-ного тельца четко варьирует в соответствии с изменением РаС()2 (рис. 16-4Б), дополнительная афферентная активность в ответ на метаболические или дыхательные стимулы не влияет на степень повышения минутной вентиляции, вызываемой центральными хеморецепторами. Более того, денервация каротидного тельца снижает вентиляторный ответ на повышение РаС()2 не менее чем на 20 %. Это служит дополнительным доказательством того, что каротидное тельце играет небольшую роль в вентиляторной реакции на ацидоз.
|
|
|
