 |


Движение с помощью ресничек и жгутиков
|
|
|
|
ГЛАВА 1. СОКРАТИТЕЛЬНЫЙ АППАРАТ БЕСПОЗВОНОЧНЫХ
Накопленный к настоящему времени большой сравнительно-физиологический материал свидетельствует о том, что все основные структуры сократительного аппарата возникли уже на самых ранних этапах филогенеза. Вместе с тем нельзя отрицать прогрессивную эволюцию двигательной функции, которая не всегда шла прямолинейным восходящим путем. Возникновение различных форм сократительного аппарата в некоторых случаях определялось не только уровнем организации животных, но и средой обитания, конкретными функциональными задачами, механизмами использования той или иной двигательной системы. Развитие сократительного аппарата многоклеточных животных происходило параллельно и взаимосвязанно с эволюцией других систем организма, в первую очередь нервной и опорной. Однако различные формы движения имеют место и в донервный период развития у одноклеточных организмов, которые перемещаются токами цитоплазмы или благодаря биению ресничек и жгутиков. В соответствии с механизмом локомоции в настоящее время различают три основных типа движения: амебоидное, ресничное и мышечное.
Амебоидное движение
Амебоидное движение, распространенное у простейших, получило свое название от способа перемещения одноклеточного — амебы (класс Rhizopoda — корненожки). Этот вид локомоции связан с токами цитоплазмы, существенным изменением формы клетки и образованием временных органелл движения — псевдоподий.
Цитоплазма амебы состоит из наружного вязкого слоя — эктоплазмы (плазмагель) и внутреннего более жидкого слоя — эндоплазмы (плазмазоль). Эндоплазма находится в постоянном движении, переливаясь к переднему (по направлению движения амебы) концу клетки (рис. 1). Ток эндоплазмы оказывает давление на клеточную оболочку — плазмолемму, растягивает ее и способствует образованию на переднем конце клетки псевдоподии. В апикальной зоне псевдоподий плазмазоль быстро переходит в плазмагель, образующий плотный гиалиновый колпачок.
|
|
|
От переднего конца клетки эндоплазма перетекает к боковой поверхности плазмолеммы и здесь изменяет свое коллоидное состояние, переходя в плазмагель. На заднем конце амебы плазмагель превращается в плазмазоль, и далее весь цикл повторяется.
Согласно современным представлениям, механизм скольжения плазмазоля по поверхности плазмагеля в своей основе сходен с механизмом мышечного сокращения. Эта гипотеза базируется на том факте, что во всех эукариотических клетках имеются белки актин и миозин, способные к обратимому связыванию друг с другом. Актины различных клеток идентичны по составу и образуют в цитоплазме тонкие транспортные нити (микрофиламенты). Миозины из различных источников более специфичны по своему составу, однако все они могут обратимо присоединяться к актиновым нитям и катализировать гидролиз аденозинтрифосфата (АТФ), который обеспечивает энергией процесс скольжения. Реакция между цитоплазматическим актином и миозином, как и в случае мышечного сокращения, может происходить только при наличии в клетке адекватных концентраций ионов кальция. Так, введение в клетку веществ, связывающих Са2+ приводит к прекращению амебоидного движения.
Исследование ультраструктуры контрактильных систем простейших показало, что обязательными элементами этих систем являются микрофиламенты и цитоплазматические микротрубочки. Л. Н. Серавин предложил различать два типа сократительных систем одноклеточных животных в зависимости от наличия в цитоплазме указанных выше элементов. К первому типу он относит однокомпонентные системы, состоящие только из микротрубочек или только из микрофиламентов. Второй тип образуют двухкомпонентные системы, построенные из ультраструктур обоего рода или же из микрофиламентов двух видов — толстых и тонких.
|
|
|
У простейших широко распространены оба типа контрактильных систем. Например, в эктоплазме гигантской болотной амебы обнаружены и микрофиламенты, и микротрубочки, в то время как контрактильная система почвенной амебы состоит только из микрофиламентов. Такое разнообразие контрактильных систем одноклеточных животных, по мнению Л. Н. Серавина, позволяет предположить относительно независимое их возникновение в процессе эволюции. Механизм амебоидного движения при наличии двухкомпонентной контрактильной системы может быть обусловлен скольжением несущих сократительные белки ультраструктур друг относительно друга.

Контрактильные системы простейших изучены еще недостаточно, однако в настоящее время для их исследования открываются широкие возможности.
Движение с помощью ресничек и жгутиков
Многие организмы, имеющие малые размеры тела, передвигаются в окружающей их жидкой среде с помощью ресничек или жгутиков. К таким животным относятся инфузории, жгутиконосцы, гребневики, мелкие ресничные черви.
Реснички довольно часто встречаются и внутри тела многоклеточных: в эпителиальных тканях дыхательных, выделительных и репродуктивных органов. Независимо от положения вне или внутри организма реснички и жгутики имеют сходную структуру. Как показали результаты электронно-микроскопических исследований, каждая ресничка состоит из двух центральных нитей (фибрилл), окруженных девятью периферическими (рис. 2). Эту комбинацию фибрилл (9+2) можно считать универсальной, так как она встречается почти у всех групп животных от простейших до млекопитающих.

Анализ структуры и функции ресничек показал, что все периферические фибриллы представляют собой парные образования, формируемые из микротрубочек А и В. Обе микротрубочки состоят из сократительного белка тубулина. Кроме того, каждая А-микротрубочка увенчана двумя выступами, образованными другим сократительным белком — динеином. Этот белок обладает способностью катализировать расщепление АТФ и обратимо прикрепляться к соседним тубулиновым микротрубочкам. Центральные фибриллы соединяются с А-микротрубочками при помощи радиальных перекладин (см. рис. 2). В основании каждой реснички лежит базальное тельце, которое, по всей вероятности, служит матрицей для сборки микротрубочек.
|
|
|
Согласно современным данным, механизм перемещения ресничек сходен с механизмом актин-миозинового взаимодействия в скелетной мышце. Полагают, что движение ресничек происходит в результате присоединения динеиновых выступов А-микротрубочек к соседним В-микротрубочкам и их взаимного скольжения за счет энергии гидролитического расщепления АТФ. Движение реснички начинается со смещения пяти периферических фибрилл. Четыре фибриллы противоположной стороны реснички включаются в работу позже, и благодаря этому процессу ресничка медленно возвращается в исходное положение. Установлено, что направление движения простейших определяется изменением внутриклеточной концентрации ионов кальция. Так, например, инфузории, встречающие на своем пути препятствие, начинают пятиться, что обусловлено открытием кальциевых каналов клеточной мембраны и притоком ионов кальция в цитоплазму.
Биение ресничек происходит в определенной последовательности, при этом колебания ресничек, расположенных в соседних рядах, незначительно сдвинуты по фазе, что создает картину так называемых метахрональных волн, пробегающих вдоль тела в одном направлении.
Координирующую роль в формировании метахрональных волн гипотетически приписывают фибриллярной сети. Эта сеть представляет собой систему тонких аргентофильных нитей, соединяющих между собой базальные тельца. Возможно, в донервный период у одноклеточных фибриллярная сеть берет на себя функцию распространения возбуждения, однако проводить аналогию с нервной системой нельзя. В регуляции автоматических движений отдельной реснички у простейших принимают участие базальные тельца и поверхностная мембрана.
|
|
|
1.3. Мышечные формы движения
 |
1.3.1. Сократительный аппарат и локомоция кишечнополостных В ходе филогенеза иннервация сократительного аппарата, впервые появляется у кишечнополостных. Он представлен миофибриллами, которые лежат в основании полифункциональных эпителиально-мышечных клеток. Эти клетки выполняют одновременно как покровную, так и двигательную функции (рис. 3).
В эпидермальной сети миофибриллы ориентированы вдоль, а в гастродермальной поперек оси тела животного. В местах контактов мембраны соседних эпителиально-мышечных клеток образуют поперечно исчерченные утолщения — десмосомы.
По своим функциональным свойствам лишенные поперечной исчерченности миофибриллы кишечнополостных напоминают гладкие мышцы более высокоорганизованных животных. Миофибриллы обладают большой пластичностью и не имеют определенной длины в состоянии покоя. Их сокращение в ответ на одиночный стимул характеризуется относительно быстрым нарастанием и медленным (до 1 мин) расслаблением (спадом).
Двигательный аппарат гидроидных медуз, сифонофор и сцифомедуз наряду с гладкими содержит поперечнополосатые волокна. Поперечнополосатые волокна эпителиально-мышечных клеток нижней стороны зонта у медуз формируют мощную кольцевую мышцу. В результате резкого сокращения этой мышцы происходит выталкивание воды из-под зонта, что обеспечивает поступательное движение животного по реактивному принципу. Упругое сопротивление в этой механической системе создает эластичная мезоглея, которая благодаря своей упругости вызывает вновь расправление зонта медузы. Саркомеры поперечнополосатых миофибрилл по своему строению схожи с саркомерами скелетных мышц других животных. Здесь обнаруживаются четко выраженные темные анизотропные и светлые изотропные диски, образованные толстыми, миозиновыми, и тонкими, актиновыми, протофибриллами. Вместе с тем, некоторый примитивизм организации проявляется в меньшей выраженности ограничивающих саркомер Z-мембран и недоразвитости саркоплазматического ретикулума, который представлен небольшим количеством везикул.
В отличие от гладкомышечных поперечнополосатые волокна медузы имеют определенную длину в покое. Длительность одиночного сокращения кольцевой мышцы медузы не превышает 2 с. Однако скорость сокращения мышц у кишечнополостных не всегда коррелирует с наличием поперечной исчерченности. Так, например, гладко-мышечный мезентериальный ретрактор актинии Metridium способен давать быстрые сокращения длительностью 0.1 с. В то же время, поперечнополосатые мышечные волокна могут сокращаться как быстро, так и медленно.
|
|
|
Выше отмечалось, что сократительный аппарат кишечнополостных активируется нервной сетью. Распространение возбуждения от рецепторов к мышце может осуществляться как по многозвенным путям с низкой скоростью (0,12-0,15 м/с), так и по прямым трактам без переключений, со скоростью проведения порядка 1,2 м/с.
Нервная сеть кишечнополостных играет существенную роль в общей координации функций двигательного аппарата, который может осуществлять не только локомоцию, но и захват пищи, защитные реакции. Так, например, пресноводной гидры тактильные раздражения вызывают втягивание щупалец и ротового диска. С тем же успехом при изменении общей формы тела гидра может осуществлять шагающие движения, попеременно прикрепляясь к грунту то оральным концом, то подошвой.
Таким образом, локомоторный аппарат оказывается тесно связанным с особенностями биологии и жизненных отправлений животного, которые реализуются при участии нервной системы.
1.3.2. Мышцы и способы передвижения червей Плоские черви, Покровы и сократительный аппарат плоских червей, срастаясь друг с другом, формируют комплексное образование, известное под названием кожно-мускулъного мешка. Он состоит из наружных кольцевых и внутренних продольных мышц, которые дополняются диагональными мышечными волокнами, пронизывающими мезенхиму в дорсовентральном направлении.
Продольные и кольцевые мышцы — это функциональные антагонисты, у свободноживущих ресничных червей (турбеллярий) участвуют в процессах ползания и плавания. Вместе с тем мелкие формы турбеллярий передвигаются главным образом благодаря работе ресничек кожного эпителия. Работа косых мышечных волокон приводит к изменению формы тела ресничных червей — сплющиванию его в дорсовентральном направлении.
Ультраструктурная организация мышечных волокон плоских червей наиболее детально изучена у пресноводной планарии Dugesia tigrina. Мышечные клетки этих животных веретеновидной формы и довольно мелкие (диаметр 1,3-3,5 мкм). Под складчатой клеточной мембраной в саркоплазме находится сократительный аппарат, состоящий из толстых и тонких протофибрилл. Тонкие протофибриллы располагаются между толстыми без всякой системы, и анизотропные и изотропные диски в саркомере не обнаруживаются.
 |
В примембранном слое саркоплазмы локализованы везикулы и цистерны саркоплазматического ретикулума. Иннервация мышечных волокон осуществляется нервными волокнами, терминали которых содержат мелкие овальные и большие гранулированные синаптические пузырьки.
Данные о функциях сократительного аппарата плоских червей весьма немногочисленны. Так, например, установлено возбуждающее действие ионов кальция на спонтанные сокращения продольных мышц у турбеллярий. Отмечается также активирующий эффект от аппликации на мышцу таких медиаторных веществ, как адреналин, норадреналин и серотонин.
Круглые черви. Классический пример упрощения организации при переходе к паразитизму дает сократительный аппарат круглых червей. Традиционным объектом при исследовании их сократительного аппарата являются мышечные клетки аскариды.
Кожно-мускульный мешок этого животного состоит из одного слоя продольных мышечных волокон, которые позволяют телу только изгибаться в различных направлениях. Перистальтические движения у аскариды отсутствуют. Мышечные клетки паразита характеризуются полифункциональностью. Каждая крупная клетка состоит из трех функционально дифференцированных частей: трофической — перинуклеарной, сократительной и иннервационной (рис. 4).
Шарообразная перинуклеарная часть (брюшко) содержит ядро, митохондрии, гранулы гликогена. Лентообразная сократительная часть имеет косую исчерченность и состоит из толстых (23 нм) и тонких (12 нм) протофибрилл, между которыми имеются тонкие поперечные мостики. В отличие от поперечнополосатых мышечных волокон в мышечных клетках аскариды толстые протофибриллы расположены в лестничном порядке. Они лежат не одна под другой, а смещены каждая последующая относительно предыдущей примерно на 10% своей длины.
Иннервационная часть клетки представляет собой трубчатый отросток, который несколькими пальцеобразными выростами вплотную подходит к нервному стволу. Выросты образуют синаптические контакты с волокнами нервного ствола. В области нервно-мышечного контакта в нервных волокнах обнаруживаются скопления везикул диаметром порядка 50 нм. Таким образом, иннервационный отросток мышечной клетки аскариды берет на себя проводниковую функцию, и в какой-то мере соответствует периферическим моторным нервам более высоко организованных животных.
Иннервационные отростки соседних мышечных клеток соприкасаются между собой и образуют плотные контакты, через которые осуществляется электротоническое взаимодействие. Эта особенность превращает мышцу аскариды в функциональный синцитий, идентичный синцитию гладких мышц у позвоночных животных.
Электрофизиологические эксперименты показывают, что мембранный потенциал мышечных клеток аскариды невелик (в среднем 30 мВ) и уменьшается при возрастании наружной концентрации ионов хлора. Низкий уровень мембранного потенциала коррелирует со способностью клеток к миогенному автоматизму, что проявляется в периодической генерации спонтанных потенциалов действия амплитудой до 10 мВ. Потенциалы действия возникают в иннервационных отростках и далее проводятся к сократительным элементам клеток. В месте генерации спайков локализованы ацетилхолиновые рецепторы, которые при обратимом связывании с ацетилхолином стимулируют процессы изменения проницаемости и деполяризации мембраны мышечной клетки. Как следствие этих процессов, при ионофоретической аппликации медиатора наблюдается сокращение соматических мышц аскариды.
Антагонистом ацетилхолина для мышечных клеток аскариды выступает гаммааминомасляная кислота, микроаппликация которой вызывает гиперполяризаию мембраны сократительной клетки и подавление спонтанной генерации потенциалов действия.
В условиях острого эксперимента эффекты, идентичные описанным выше, могут быть получены при электрическом раздражении нервного ствола. Характер ответа (активация или торможение) мышечной клетки определяется положением стимулирующих электродов и силой раздражения.
Эти факты позволяют допустить наличие двойной иннервации мышечных клеток аскариды — возбуждающей и тормозной. Этот принцип является общим для функционирования соматической мускулатуры многих беспозвоночных животных.
Кольчатые черви. Кожно-мускульный мешок малощетинковых кольчецов образован двумя слоями мышц-антагонистов: наружным кольцевым и внутренним продолъным Кольцевая мускулатура земляных червей состоит из мышечных пучков, разделенных междусегментными перегородками — септами. Продольные мышцы, напротив, тянутся через несколько сегментов и присоединяются одним концом к септам, а другим свободно оканчиваются в соединительной ткани.
Координированная деятельность обеих групп мышц лежит в основе локомоции кольчецов, которая обусловлена перистальтической волной сокращений, распространяющихся вдоль тела животного. Как правило, сокращение кольцевых мышц начинается на переднем конце тела и вызывает давление на целомическую жидкость, которая растягивает продольные мышцы и изменяет форму головных сегментов. Передний конец тела червя становится длиннее и тоньше и продвигается таким образом вперед. Щетинки на переднем конце тела втягиваются и не препятствуют поступательному движению червя. В это время на заднем конце тела сокращаются продольные мышцы, объем задних сегментов увеличивается, выдвигаются щетинки. В результате задний конец тела фиксируется относительно окружающей почвы, и животное отталкивается от нее. Последующее укорочение переднего конца тела приводит к подтягиванию задних сегментов к передним, и далее весь цикл волнообразных перистальтических сокращений повторяется заново.
У многощетинковых червей (полихет) кольцевая мускулатура развита слабее, чем у олигохет, и осуществляет главным образом перемещение полых выступов тела параподий. Рабочее движение параподий происходит в результате повышения гидростатического давления в целоме, как следствие сокращения мышц стенки тела. Параподия цепляется за субстрат и оттаткивается от него. Далее, после сокращения сегментарных кольцевых мышц, параподия втягивается и переносится вперед. Эти шагающие движения параподий позволяют полихете ползать по субстрату. Работа всех параподий сопряжена с возникновением перистальтических волн, идущих от хвостового конца тела к голове; при этом перемещения, параподий правой и левой стороны реципрокны — движению параподий одной стороны тела вперед соответствует перемещение симметричных им параподий другой стороны назад. Плавание у полихет осуществляется в результате сокращения правой и левой продольных мышц, что создает волны горизонтальных изгибов тела и веслообразные перемещения параподий.
Исследование ультраструктурной организации мышечных волокон кольчатых червей показало, что они представлены одноядерными клетками с периферическим расположением ядра и саркоплазмы. Центральную часть клетки занимает сократительный аппарат, состоящий из тонких и толстых протофибрилл. Протофибриллы организованы в анизотропные и изотропные диски, ориентированные под углом к продольной оси волокна. Соседние ряды толстых протофибрилл смещены друг относительно друга примерно на 10% своей длины.

Система внутренних полостей и трубочек внутри волокна формирует саркоплазматический ретикулум. Выпячивания клеточной мембраны, дающие начало поперечным Т-трубочкам, в мышечных волокнах земляных червей не обнаружены.
В основе сокращения косо исчерченных мышц кольчатых червей лежит механизм скольжения тонких, актиновых, протофибрилл относительно толстых миозиновых. Миозин толстых протофибрилл состоит из двух фракций А и В, причем миозин В обладает АТФ-азной активностью. Энзиматическое расщепление АТФ в мышцах аннелид активируется ионами кальция, который содержится в саркоплазматическом ретикулуме. Экспериментально установлено, что фрагменты саркоплазматического ретикулума дождевого червя способны активно поглощать наружный кальций в присутствии ионов магния и АТФ. Таким образом, рабочий механизм сократительного аппарата кольчецов весьма сходен с механизмом скольжения протофибрилл в поперечнополосатых мышцах позвоночных. Однако, по мнению Д. Розенблюта, своеобразие косоисчерченных мышц проявляется в том, что помимо скольжения тонких протофибрилл относительно толстых здесь одновременно имеет место смещение толстых филаментов друг относительно друга. Природа данного феномена пока неясна, вместе с тем очевидно, что этот дополнительный механизм увеличивает степень сокращения и растяжимость мышечного волокна кольчецов.
Соматическая мускулатура кольчатых червей так же, как и у других беспозвоночных, имеет множественную (полинейрональную) иннервацию. Аксоны сегментарных нервов, по данным Д. Розенблюта, образуют на сократительных волокнах дождевых червей нервно-мышечные синапсы двух типов.
Первый их них содержит в пресинаптическом окончании сферические пузырьки диаметром до 50 нм.
Результаты физиологических экспериментов свидетельствуют, что медиатором возбуждающего, действия у кольчецов является ацетилхолин. Так, например, при раздражении моторного нерва пиявки он появляется в перфузате, омывающем мышцу. Ацетилхолин и холинэстераза обнаружены в гомогенатах мышечной ткани олигохет, полихет и пиявок. Сокращение мышцы, вызванное стимуляцией нерва, у кольчецов усиливается при действии эзерина и блокируется такими холинолитиками, как кураре и атропин. Все эти факты подтверждают медиаторную роль ацетилхолина в мионевральных синапсах кольчецов.
Нервно-мышечный синапс второго типа характеризуется наличием электронно-плотных, гранулированных везикул (диаметром до 100 нм) в пресинаптическом окончании. Рецепторы на постсинаптической мембране синапсов данного вида не сконцентрированы в виде гранул. Наблюдается довольно плотное прилегание нервных окончаний к мембране волокна. Перечисленные выше морфологические особенности позволяют отнести синапсы данного типа к адренергическим, которые, по всей вероятности, участвуют в передаче тормозных влияний на сократительный аппарат.
Однако число претендентов на роль тормозных медиаторов в мионевральных синапсах кольчецов не ограничивается катехоламинами. Обнаружено гиперполяризующее действие гамма-аминомасляной кислоты на мембрану мышечного волокна полихет и олигохет. Микроионофоретическая аппликация 5-гидрокситриптамина также вызывает тормозный эффект и подавляет возникновение потенциалов действия в мышце пиявки. Некоторый разброс данных говорит о необходимости дополнительных, комплексных исследований механизма периферического торможения у кольчатых червей.
Существенную роль в регуляции местных сократительных реакций у кольчецов играет субэпителиальная нервная сеть. По данным О. Ф. Давида, у дождевых червей спонтанные периодические сокращения мышц стенки тела сохраняются после удаления брюшной нервной цепочки. Таким образом, наблюдаемая периодическая мышечная активность не инспирирована нервными центрами, а имеет периферический, нейрогенный механизм запуска, основанный на наличии связей между афферентными и эфферентными элементами субэпителиальной нервной сети. Нанесение на поверхность кутикулы различных анестетиков (новокаин, кокаин) обратимо выключает функцию периферических чувствительных клеток и подавляет спонтанные сокращения мышц. Вместе с тем центральная нервная система дождевых червей оказывает на периферический механизм запуска движений регулирующее воздействие, изменяя общий уровень электрической активности мышечных волокон стенки тела дождевого червя.
У полихет, пиявок, ведущих более подвижный образ жизни, удаление брюшной нервной цепочки полностью выключает спонтанные периодические сокращения мышц. Этот факт позволяет заключить, что у данных животных доминирует центральный механизм запуска ритмической активности сократительного аппарата. Очевидно, поведенческие особенности различных представителей кольчатых червей определяют больший или меньший вклад центральных структур в механизм запуска периодической сократительной активности. Вместе с тем подобное заключение справедливо только по отношению к наиболее простым видам моторных актов. При осуществлении сложных локомоторных реакций (повороты, реакции защитного характера) центральные воздействия нельзя сводить только к регулирующим. Естественно, что в этих случаях будет реализована и пусковая функция центральной нервной системы.
1.3.3. Сократительный аппарат моллюсков. В пределах обширного типа моллюсков встречаются самые разнообразные способы локомоции: ползание, ундулирующие движения, плавание, реактивное движение. Степень дифференцированности сократительного аппарата в различных таксономических группах этого типа беспозвоночных коррелирует со скоростью их передвижения и его маневренностью. У большинства "тихоходных" форм (брюхоногие, пластинчатожаберные моллюски) мышцы слабо дифференцированы и лишь немногие из них (аддукторы створок, мышцы сифонов, ретракторы биссуса и некоторые другие) представляют собой обособленные образования. В то же время, у быстрых и маневренных головоногих моллюсков индивидуализация отдельных мышц достигает значительной степени.
Весьма разнообразны мышцы моллюсков и по характеру организации самого сократительного аппарата, который может быть представлен как случайно расположенными протофибриллами (гладкие мышцы), так и четко структурированными саркомерами (поперечнополосатые мышцы). По этому признаку в пределах типа моллюсков можно выделить пять категорий мышечных волокон.
Наиболее просто организованы 1. гладкомышечные волокна, не содержащие парамиозина. Как правило, они тонкие, содержат удлиненное ядро и продольные ряды гранул. В расположении протофибрилл гладкомышечного волокна отсутствует какая-либо регулярность. Средний диаметр толстых, миозиновых, протофибрилл в ретракторе глотки улитки 25 нм, толщина тонких, актиновых, протофибрилл не превышает 5 нм.
Гладкие непарамиозиновыеволокна формируют такие мышцы, как ретрактор глотки улитки, запирательные мышцы некоторых брюхоногих моллюсков и др.
Следующий тип мышечных волокон — это 2. гладкомышечные волокна, содержащие парамиозин. Ультраструктурная организация волокон этого типа более упорядоченная, чем у волокон, рассмотренных выше. Саркоплазма волокон парамиозинового типа содержит большое количество гранул гликогена и периферически расположенные везикулы (рис. 6). Собственно сократительный аппарат представлен тонкими и толстыми протофибриллами, ориентированными по длине волокна. Тонкие, актиновые, протофибриллы имеют толщину порядка 5 нм и располагаются орбитально вокруг толстых. Толстые протофибриллы диаметром 30-150 нм и длиной до 30 мкм состоят из сократительных белков миозина и парамиозина. Как правило, миозин расположен на периферии толстой протофибриллы, а парамиозин занимает центральное положение. Содержание парамиозина в этих гладкомышечных волокнах велико и составляет около 32% общего количества белка.
Оба типа протофибрилл соединяются друг с другом поперечными мостиками, В саркоплазме волокон парамиозинового типа можно различить плотные тела, возле которых тонких протофибрилл больше. Существует мнение, что плотные тела гомологичны Z-линиям поперечнополосатых мышц. Характерной особенностью гладких парамиозиновых волокон является способность длительно поддерживать тоническое напряжение; Эти волокна обычно включены в мощные мышцы-аддукторы, которые закрывают раковину, в ретракторы биссусных нитей, фиксирующих моллюска на субстрате.
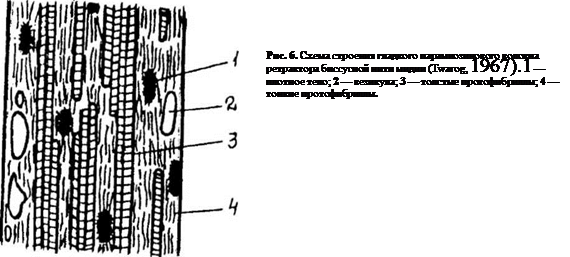
3. Поперечнополосатые мышечные волокна обнаружены в фазной части запирательных мышц двустворчатых моллюсков семейства гребешков (Pectinidae). Эти волокна имеют лентовидную форму и отличаются малой толщиной — около 5 мкм. Каждое волокно разделяется Z-линиями на саркомеры, длина которых составляет примерно 3 мкм. В пределах саркомера четко выражены анизотропные диски, содержащие толстые (до 30-40 нм) и тонкие (около 5 нм) протофибриллы. В целом, структура данного типа волокон обнаруживает большое сходство с поперечнополосатыми волокнами позвоночных животных.
4. Косоисчерченные мышечные волокна по скорости сокращения несколько уступают поперечнополосатым и встречаются в фазных аддукторах ряда представителей пластинчатожаберных, двустворчатых моллюсков (устрицы, беззубки и др.). Специфичным для этих волокон является расположение А- и I- дисков под малым углом (около 10°) к продольной оси волокна. Так же, как и у мышечных волокон червей, косая исчерченность обусловлена смещением толстых протофибрилл друг относительно друга. Другая особенность косоисчерченных мышечных волокон моллюсков состоит в отсутствии типичных Z-линий. На их месте, как и в гладких мышцах, располагаются электронно-плотные тела.
Наконец, 5. спиральные мышечные волокна, которые формируют мышцы двигательного аппарата головоногих моллюсков: продольные и кольцевые мышцы мантии, мышцы воронки, присосок, хроматофорных органов. Волокна этого типа на поперечных срезах имеют округлую или овальную форму, их диаметр достигает 5-10 мкм. Сократительный аппарат волокна состоит из плотно расположенных толстых протофибрилл, между которыми вмонтированы тонкие. Протофибриллы волокна обладают спиральным ходом, что обусловливает его двойную косую исчерченность при наблюдении в световом микроскопе. Величина угла, образуемого спиралью с продольной осью волокна, зависит от степени растяжения мышечного препарата.
Несмотря на такое разнообразие типов сократительного аппарата, мышцы моллюсков обладают целым рядом общих особенностей, касающихся, в первую очередь, характера их сокращения и иннервации.
Как правило, мышцы моллюсков (за исключением некоторых мышц головоногих) являются относительно медленными. Так, например, время достижения максимального напряжения при одиночном сокращении ретрактора биссусной нити у мидии составляет 1-1,5 с, тогда как для максимального напряжения портняжной мышцы лягушки требуется всего 0,03 c.
Характерная особенность мышц моллюсков высокая растяжимость и пластичность — способность сохранять остаточное удлинение и остаточное укорочение. Активная сократительная сила зависит от исходной длины мышцы у моллюсков больше, чем в мышцах позвоночных животных. Эти особенности обусловлены экологией моллюсков, длина мышц тела которых обычно подвергается значительным изменениям из-за отсутствия скелета и жесткого прикрепления.
Некоторые мышцы моллюсков, в первую очередь парамиозиновые, обладают значительной абсолютной силой сокращения, которая достигает 8-9 кг/см2. Для сравнения следует отметить, что среднее значение абсолютной силы сокращения мышц млекопитающих составляет 3-4 кг/см2.
Исследование иннервации мышц моллюсков наталкивается на значительные методические трудности, так как тонкие концевые разветвления нервов прокрашиваются довольно плохо. Однако электронно-микроскопическими методами у брюхоногих и пластинчатожаберных моллюсков обнаружены нервные стволы, которые идут параллельно мышечным волокнам. Их терминальные ветвления образуют на поверхности мышцы сложные сплетения. Тончайшие терминали этих сплетений (от 0,1 до 1 мкм) формируют нервно-мышечные контакты, ширина синаптической щели которых достигает 15 нм. В местах контактов мембран нервного и мышечного волокон имеются специфические уплотнения. Нервная терминаль содержит синаптические пузырьки и митохондрии. Все эти признаки характерны для мионевральных синапсов не только моллюсков, но и других животных.
Установлено, что иннервация мышц моллюсков мультитерминальная. т. е. одно мышечное волокно снабжено несколькими нервными окончаниями. Данные физиологических экспериментов позволяют говорить о наличии у моллюсков двух систем иннервации — быстрой и медленной. Так, например, циркулярная мышца мантии головоногих моллюсков обладает быстрой иннервацией, осуществляемой по гигантским (диаметр 800 мкм) аксонам звездчатого ганглия. Возбуждение по этим аксонам проводится с большой скоростью (до 25 м/с) и вызывает быстрое и одновременное сокращение множества мышечных волокон циркулярной мышцы. Синхронное сокращение большинства волокон обеспечивает мгновенный выброс воды из сифона и реактивное движение животного.
Медленная иннервация циркулярной мышцы обеспечивается системой тонких аксонов, раздражение которых вызывает градуальное сокращение. Данное сокращение лежит в основе дыхательных движений мантии, не связанных с локомоцией.
Результаты физиологических опытов дают основание думать, что наряду с возбуждающей иннервацией в мышцах моллюсков существует и тормозная. К таким экспериментам относится раздражение периферического конца педального нерва улитки, которое вызывает расслабление мышц изолированной ноги. Однако морфологическая основа тормозящих воздействий исследована пока недостаточно. Остается дискуссионным и вопрос о медиаторах тормозных и возбуждающих периферических нервов моллюсков. Возможным претендентом на роль медиатора в мионевральных синапсах моллюсков является серотонин, который в низких концентрациях воспроизводит тормозный эффект. Вместе с тем серотонин оказывает прямое действие на сократительный аппарат мышцы, изменяя внутриклеточную концентрацию кальция, т. е. не может считаться в данном случае классическим медиатором. Большинство исследователей считает, что мышцы моллюсков возбуждаются холинергическими нервами, причем холинорецепторы этих мышц являются никотинчувствительными. Однако имеются факты, которые не укладываются в данную концепцию. Так, например, быстрый аддуктор моллюсков семейства гребешков не обнаруживает чувствительности к ацетилхолину и к таким холиномиметикам, как арсколин и пилокарпин. Некоторые авторы допускают, что возбуждающим медиатором мионеврального синапса ретрактора глотки улитки является глутамат. Очевидно, что для окончательного вывода по данному вопросу еще необходимы фактическое подкрепление и расширение круга экспериментальных объектов.
К сожалению, до сих пор остается спорным и вопрос о механизмах запирательного тонуса гладких парамиозиновых мышц моллюсков. Многолетние исследования привели к созданию нескольких гипотез, объясняющих причины длительного сокращения мышц-аддукторов. Согласно одной из них, мышцы парамиозинового типа имеют двойную иннервацию — фазную и тоническую. По ходу фазных нервов синаптических переключений мало и их возбуждение вызывает быстрые сокращения мышцы. Тонические нервы, напротив, контактируют с мышцей не пря
|
|
|
