 |


Межвидовая конкуренция в природе
|
|
|
|
 В последние два десятилетия в экологической литературе ведутся ожесточенные споры по поводу того, какова роль конкуренции в ограничении распространения и динамики природных популяций разных видов, а, следовательно, и в определении структуры сообществ. По мнению одних исследователей, популяции, входящие в естественные сообщества, достаточно строго контролируются системой конкурентных отношений, иногда, правда, модифицируемых воздействием хищников. Другие же полагают, что конкуренция между представителями разных видов наблюдается в природе только эпизодически, а популяции, в большинстве своем, будучи ограничены другими факторами, как правило, не достигают тех плотностей, при которых конкурентные отношения становятся определяющими. Существует также и не лишенная оснований компромиссная точка зрения, предполагающая наличие некоторого континуума реальных природных сообществ, на одном конце которого — сообщества, стабильные во времени, богатые, или, точнее, насыщенные видами, жестко контролируемые биотическими взаимодействиями, а на другом—сообщества нестабильные (в большинстве случаев из-за того, что не стабильны абиотические условия в их местообитаниях), не насыщенные видами (т, е. допускающие вселение новых видов) и контролируемые, как правило, плохо предсказуемыми изменениями внешних условий.
В последние два десятилетия в экологической литературе ведутся ожесточенные споры по поводу того, какова роль конкуренции в ограничении распространения и динамики природных популяций разных видов, а, следовательно, и в определении структуры сообществ. По мнению одних исследователей, популяции, входящие в естественные сообщества, достаточно строго контролируются системой конкурентных отношений, иногда, правда, модифицируемых воздействием хищников. Другие же полагают, что конкуренция между представителями разных видов наблюдается в природе только эпизодически, а популяции, в большинстве своем, будучи ограничены другими факторами, как правило, не достигают тех плотностей, при которых конкурентные отношения становятся определяющими. Существует также и не лишенная оснований компромиссная точка зрения, предполагающая наличие некоторого континуума реальных природных сообществ, на одном конце которого — сообщества, стабильные во времени, богатые, или, точнее, насыщенные видами, жестко контролируемые биотическими взаимодействиями, а на другом—сообщества нестабильные (в большинстве случаев из-за того, что не стабильны абиотические условия в их местообитаниях), не насыщенные видами (т, е. допускающие вселение новых видов) и контролируемые, как правило, плохо предсказуемыми изменениями внешних условий.
 Получить прямые доказательства важности роли конкуренции в определении динамики и распределения популяций в природе весьма нелегко. Обычно мы можем судить об этом только на основании косвенных свидетельств, но заметим, что сам по себе косвенный характер тех или иных свидетельств не должен служить основанием для их игнорирования. В тех случаях, когда ряд независимо полученных косвенных свидетельств выстраивается в логически обоснованную и не противоречащую здравому смыслу схему, не следует отвергать эту схему на том лишь основании, что отсутствуют прямые доказательства. Надо также подчеркнуть, что непосредственно наблюдать в природе сам процесс конкуренции удается не так уж часто. Основная же масса имеющихся свидетельств конкуренции касается такого распределения видов относительно друг друга в пространстве или времени, которое может быть трактовано как результат конкуренции. Ниже мы приведем несколько примеров подобного распределения.
Получить прямые доказательства важности роли конкуренции в определении динамики и распределения популяций в природе весьма нелегко. Обычно мы можем судить об этом только на основании косвенных свидетельств, но заметим, что сам по себе косвенный характер тех или иных свидетельств не должен служить основанием для их игнорирования. В тех случаях, когда ряд независимо полученных косвенных свидетельств выстраивается в логически обоснованную и не противоречащую здравому смыслу схему, не следует отвергать эту схему на том лишь основании, что отсутствуют прямые доказательства. Надо также подчеркнуть, что непосредственно наблюдать в природе сам процесс конкуренции удается не так уж часто. Основная же масса имеющихся свидетельств конкуренции касается такого распределения видов относительно друг друга в пространстве или времени, которое может быть трактовано как результат конкуренции. Ниже мы приведем несколько примеров подобного распределения.
|
|
|
Исследуя изменения видового состава птиц в Перуанских Андах по мере подъема в горы, Я. Терборгх (Terborgh, 1971) обнаружил, что виды одного рода очень четко сменяют друг друга, причем границы распространения часто не связаны с вертикальной поясностью растительности, а определяются, вероятно, только конкуренцией между близкими видами. На схеме (рис. 57), заимствованной из работы Я. Терборгха видно, что чем больше видов одного рода встречено на всем обследуемом диапазоне высот, тем меньший интервал высот приходится в среднем на один вид. Так, если от высоты 1000 до высоты 3400 м встречаются два представителя одного рода, то на каждого приходится интервал в 1200 м, а если в таком же диапазоне высот обитают три вида одного рода, то на каждый вид приходится в среднем по 800 м. Подобное распределение явно указывает на конкуренцию, и оно вряд ли может быть объяснено без учета межвидовых взаимодействий (MacArthur, 1972). Важные дополнительные свидетельства наличия конкуренции в случае, описанном Я. Терборгхом, были получены при изучении вертикального распределения птиц, проведенном при участии того же автора (Terborgh, Weske, 1975) в Андах, но не на основном хребте, а на небольшом изолированном горном массиве, расположенном от него в 100 км. Число обитающих здесь видов было значительно меньше, чем на хребте, но те же самые виды встречались в большем диапазоне высот, указывая на то, что именно конкурентные отношения, а не абиотические факторы ограничивают их распространение на основном хребте.
|
|
|
Много примеров межвидовой конкуренции дает островная фауна (Майр, 1968), представители которой нередко демонстрируют взаимоисключающее распределение, хотя на материке живут бок о бок. Так, М. Радованович (Radovanovic, 1959; цит. по: Майр, 1968), изучив распространение ящериц рода Lacerta на 46 островах Средиземного моря около побережья Югославии, выяснил, что на 28 островах встречалась только Lacerta melisellensis, а на остальных — только Lacerta sicula. Нет ни одного острова, на котором оба вида обитали бы вместе.
В более редких случаях исследователи могли непосредственно наблюдать расширение области распространения одного вида, сопровождающееся исчезновением или сокращением численности в этой области другого вида, являющегося его потенциальным конкурентом. Так, с конца XIX вплоть до середины XX в. в Европе было замечено резкое сокращение ареала широкопалого рака (Astacus astacus) и соответствующее расширение на северо-запад ареала близкого вида — длиннопалого рака (Astacus lepiodactylus), захватившего весь Волжский бассейн, а затем проникшего в бассейн Невы и Северского Донца (Бирштейн, Виноградов, 1934). В настоящее время оба вида встречаются в Прибалтике и Белоруссии, однако, случаи их нахождения в одном водоеме очень редки (Цукерзис, 1970). Механизм вытеснения одного вида другим не ясен, за исключением тех немногих случаев, когда длиннопалый рак был специально запущен в те водоемы, где широкопалый погиб при эпизоотии «рачьей чумы» — грибкового заболевания, способного полностью уничтожить популяцию речных раков. Вполне вероятно, что успешному расширению ареала A. lepiodactylus способствовало и то, что по сравнению с A. astacus он быстрее растет, отличается большей плодовитостью и способностью питаться круглосуточно, а не только ночью, как широкопалый рак.
На территории Британских островов было замечено резкое сокращение ареала обыкновенной белки (Sciurus vulgaris) после завоза из Северной Америки близкого к ней вида Каролинской белки (Sciurus carolinensis), хотя природа конкурентного вытеснения осталась неизвестной. Виды, обитающие на островах, особенно страдают от вселенцев с материка, которые, как правило, оказываются более конкурентоспособными. Как отмечает Э. Майр (1968), большинство видов птиц, исчезнувших за последние 200 лет, были островными.
|
|
|
Очевидно, что увеличение области распространения одного вида, совпадающее с одновременным сокращением области распространения другого экологически близкого вида, вовсе не обязательно должно быть следствием конкуренции. На подобный сдвиг границ зон обитания могут влиять также иные биотические факторы, например деятельность хищников, доступность кормовых объектов или изменение абиотических условии. Так, в качестве примера конкурентного вытеснения рассматривалось ранее изменение распространения на Ньюфаундленде двух видов зайцев: полярного зайца (Lepus arcticus) н американского зайца-беляка (Lepus americanus). Более ста лет тому назад на острове обитал только полярный заяц, который населял самые различные биотопы, как в горах, так и в лесных долинах. Завезенный на остров в конце прошлого века заяц-беляк распространялся по лесным долинам, а полярный заяц стал встречаться только в горных безлесных районах. Была предложена простая гипотеза конкурентного вытеснения одного вида другим, но затем выяснилось (Bergerud, 1967), то в исчезновении полярного зайца из лесных районов виновен хищник — рысь (Lynx lynx), численность которой резко возросла после вселения на остров зайца-беляка. Косвенным доводом в пользу того, что пресс хищников сыграл в данном случае решающую роль, служит исчезновение полярного зайца из тех районов, куда не проник заяц-беляк, но которые по характеру растительности удобны для преследования зайцев рысью. Таким образом, гипотеза конкурентного исключения в данном случае, хотя и не была отвергнута полностью, должна была уступить гипотезе, учитывающей взаимоотношения трех видов: двух потенциальных конкурентов и одного хищника.
Сосуществование конкурирующих видов. Модели динамики, определяемой концентрацией ресурсов
|
|
|
Если достоверно доказанных случаев конкурентного вытеснения одного вида другим в природных условиях весьма немного, а о значении конкуренции как фактора, определяющего динамику популяций и сообществ, ведутся нескончаемые дискуссии, то сами по себе многочисленные факты сосуществования экологически близких и потому скорее всего конкурирующих видов сомнения не вызывают. Так, выше мы уже упоминали о «планктонном парадоксе», но с не меньшим основанием можно говорить и о «парадоксе луга», поскольку ряд видов травянистых растений, ограниченных светом, влагой и одним и тем же набором элементов минерального питания, произрастают бок о бок в одном месте, хотя и находятся в конкурентных отношениях.
В принципе сосуществование конкурирующих видов (т, е. несоблюдение закона Гаузе) может быть объяснено следующими обстоятельствами: 1) популяции разных видов ограничены разными ресурсами; 2) хищник преимущественно выедает более сильного конкурента; 3) конкурентное преимущество видов изменяется в зависимости от непостоянства внешних условий (т. е. конкурентное вытеснение каждый раз не доходит до конца, сменяясь периодом, благоприятным для вида, ранее вытесняемого); 4) популяции разных видов на самом деле разделены в пространстве — времени, и то, что представляется наблюдателю одним местообитанием, с точки зрения изучаемых организмов содержит целый набор разных местообитаний.
Чтобы объяснить сосуществование видов, конкурирующих за ограниченное число ресурсов, необходимо хотя бы вкратце рассмотреть модель динамики популяций, лимитированных в своем развитии количеством имеющегося ресурса. В основе этой модели лежит уже упоминавшееся выше представление о так называемой пороговой концентрации ресурса R*, т. е. той минимальной концентрацией, при которой рождаемость точно уравновешена смертностью (см. рис. 44), а популяция сохраняет стационарную численность. Очевидно, у разных видов, зависящих от одного ресурса, значения пороговых концентраций могут не совпадать, но если ресурса в окружающей среде много, то оба вида растут с максимальными скоростями, и быстрее наращивает свою численность тот вид, у которого больше при данной концентрации разность рождаемости и смертности (т. е. величина b – d). Очевидно, однако, что в природной обстановке по мере увеличения количества организмов, потребляющих данный ресурс, концентрация его в среде снижается, а когда она достигает пороговой для данного вида организмов величины, численность популяции начинает падать. В результате конкуренции двух видов за один ресурс побеждает тот, для которого ниже пороговая концентрация ресурса[56].
|
|
|
 Теперь рассмотрим модель с двумя ресурсами, величины концентраций которых в среде R1 и R2 отложим на двух ортогональных осях (рис. 58). В координатном пространстве этих ресурсов проведем линию, соответствующую тем значениям концентраций первого и второго ресурсов, при которых популяция сохраняет свою численность постоянной (dN/Ndt = 0). Эта линия, называемая изоклиной нулевого прироста, фактически соответствует пороговым для данного вида комбинациям концентраций первого и второго ресурсов. Если точки, соответствующие наблюдаемым в среде концентрациям ресурсов, лежат от этой линии ближе к началу координат, то численность популяции при данных значениях концентраций будет падать. Если же они лежат за изоклиной, то численность популяции будет расти.
Теперь рассмотрим модель с двумя ресурсами, величины концентраций которых в среде R1 и R2 отложим на двух ортогональных осях (рис. 58). В координатном пространстве этих ресурсов проведем линию, соответствующую тем значениям концентраций первого и второго ресурсов, при которых популяция сохраняет свою численность постоянной (dN/Ndt = 0). Эта линия, называемая изоклиной нулевого прироста, фактически соответствует пороговым для данного вида комбинациям концентраций первого и второго ресурсов. Если точки, соответствующие наблюдаемым в среде концентрациям ресурсов, лежат от этой линии ближе к началу координат, то численность популяции при данных значениях концентраций будет падать. Если же они лежат за изоклиной, то численность популяции будет расти.
 Заметим, что прямой изоклина на рассматриваемом графике проведена только для простоты. Этот случай соответствует взаимозаменяемости ресурсов, т. с. возможности вида успешно существовать, потребляя только один из ресурсов или довольствуясь какой-нибудь их комбинацией. На самом деле, изоклина может быть вогнутой (комплементарность ресурсов) в тех случаях, когда, питаясь смесью разных компонентов, организм потребляет их в сумме меньше, чем при питании каждым из этих компонентов по отдельности, и выпуклой, например, при синергизме воздействия токсических веществ, потребляемых с разными пищевыми компонентами. Обратите внимание на то, что для поддержания постоянной численности одному виду (рис. 58, а) требуется гораздо больше второго ресурса, чем первого, но другой вид (рис. 58, б) может быть более эффективным потребителем именно второго ресурса, которого требуется ему соответственно меньше, чем первого.
Заметим, что прямой изоклина на рассматриваемом графике проведена только для простоты. Этот случай соответствует взаимозаменяемости ресурсов, т. с. возможности вида успешно существовать, потребляя только один из ресурсов или довольствуясь какой-нибудь их комбинацией. На самом деле, изоклина может быть вогнутой (комплементарность ресурсов) в тех случаях, когда, питаясь смесью разных компонентов, организм потребляет их в сумме меньше, чем при питании каждым из этих компонентов по отдельности, и выпуклой, например, при синергизме воздействия токсических веществ, потребляемых с разными пищевыми компонентами. Обратите внимание на то, что для поддержания постоянной численности одному виду (рис. 58, а) требуется гораздо больше второго ресурса, чем первого, но другой вид (рис. 58, б) может быть более эффективным потребителем именно второго ресурса, которого требуется ему соответственно меньше, чем первого.
Попробуем теперь на том же графике провести изоклину нулевого прироста для второго вида. Очевидно, что если изоклина вида В пройдет ближе к началу координат, чем изоклина вида А (рис. 58, б), то победителем в конкуренции будет вид В, поскольку он «доведет» концентрацию обоих ресурсов до такого низкого уровня, при котором стационарная популяция вида А существовать не сможет. Если же изоклина вида В пройдет дальше от начала координат, чем изоклина вида А, то победителем в конкуренции окажется именно вид А (рис. 58, г). Если изоклины двух видов пересекаются, то при определенном соотношении ресурсов в среде виды могут сосуществовать, а при другом — может наблюдаться вытеснение одного вида другим. Так, например, в ситуации, изображенной на рис. 58, д, при высокой концентрации второго ресурса и низкой концентрации первого конкурентное преимущество сказывается у вида А, а при высокой концентрации первого ресурса и низкой концентрации второго преимущество оказывается у вида В.

 Рассмотренный выше пример соответствовал ресурсам, полностью взаимозаменяемым. Однако для большинства организмов существует некоторое число ресурсов незаменимых. Так, например, как бы ни было хорошо обеспечено какое-либо растение азотом, оно не сможет расти и развиваться, если в его питательной среде не будет фосфора. В координатных осях двух ресурсов изоклина нулевого прироста популяции, ограниченной таким" двумя ресурсами, будет изображаться линией, изогнутой под прямым углом, т. е. так, что она оказывается состоящей из двух ветвей, параллельных осям графика (рис. 59, а). Положение каждой ветви отвечает пороговой концентрации первого или второго ресурса. Если за два незаменимых ресурса конкурируют два вида, то так же, как в случае с взаимозаменяемыми ресурсами, возможны разные варианты расположения относительно друг друга изоклин нулевого прироста этих видов. Очевидно, что в ситуации, изображенной на рис. 59, б, победителем будет вид А, а в изображенной на рис. 59, б — вид В. При пересечении изоклин (рис. 59, г) может быть достигнуто сосуществование обоих видов, так как для каждого из них лимитирующими оказываются разные ресурсы.
Рассмотренный выше пример соответствовал ресурсам, полностью взаимозаменяемым. Однако для большинства организмов существует некоторое число ресурсов незаменимых. Так, например, как бы ни было хорошо обеспечено какое-либо растение азотом, оно не сможет расти и развиваться, если в его питательной среде не будет фосфора. В координатных осях двух ресурсов изоклина нулевого прироста популяции, ограниченной таким" двумя ресурсами, будет изображаться линией, изогнутой под прямым углом, т. е. так, что она оказывается состоящей из двух ветвей, параллельных осям графика (рис. 59, а). Положение каждой ветви отвечает пороговой концентрации первого или второго ресурса. Если за два незаменимых ресурса конкурируют два вида, то так же, как в случае с взаимозаменяемыми ресурсами, возможны разные варианты расположения относительно друг друга изоклин нулевого прироста этих видов. Очевидно, что в ситуации, изображенной на рис. 59, б, победителем будет вид А, а в изображенной на рис. 59, б — вид В. При пересечении изоклин (рис. 59, г) может быть достигнуто сосуществование обоих видов, так как для каждого из них лимитирующими оказываются разные ресурсы.
 Последнему случаю есть и экспериментальное подтверждение. Так, Дэвид Тильман (Tilman, 1982), внесший большой вклад в развитие современных представлений о конкуренции за ресурсы,. провел серии экспериментов с двумя видами диатомовых планктонных водорослей Asterionella formosa и Cyclotella meneghiniaпа и на основании полученных данных построил для них изоклины нулевого прироста в зависимости от концентрации двух незаменимых ресурсов — фосфора и кремния (рис. 60).
Последнему случаю есть и экспериментальное подтверждение. Так, Дэвид Тильман (Tilman, 1982), внесший большой вклад в развитие современных представлений о конкуренции за ресурсы,. провел серии экспериментов с двумя видами диатомовых планктонных водорослей Asterionella formosa и Cyclotella meneghiniaпа и на основании полученных данных построил для них изоклины нулевого прироста в зависимости от концентрации двух незаменимых ресурсов — фосфора и кремния (рис. 60).
В рамках данной модели сравнительно легко объяснить сосуществование разных видов, если они лимитированы разными ресурсами. Однако само понятие «разные ресурсы» нуждается в уточнении. Так, наверное, все согласятся с тем, что разные виды растений для жнвотных-фитофагов могут рассматриваться как разные ресурсы. С несколько меньшими основаниями, но, видимо, можно говорить и о том, что разные части одного растения могут трактоваться как разные ресурсы. Однако количество элементов минерального питания, необходимых растениям наряду со светом и влагой очень ограничено. Во всяком случае, оно значительно меньше числа видов планктонных водорослей, обитающих в пределах небольшого объема воды (вспомните «планктонный парадокс»), или числа видов травянистых растений, произрастающих на одном лугу. Попытка объяснить сосуществование многих видов, конкурирующих за небольшое число общих ресурсов, была предпринята Д. Тильманом (Tilman, 1982). Чтобы пояснить суть его рассуждений, необходимо внести некоторые усложнения в описанную выше модель.
Начнем с того, что все предыдущие рассуждения основывались на предположении о стабильных концентрациях ресурсов. Ясно, однако, что на самом деле ресурсы, как и потребляющие их популяции, находятся в постоянной динамике пли, во всяком случае, в состоянии динамического равновесия, при котором потребление ресурса уравновешивается притоком его в среду. Если мы представим себе, что потребителей можно изъять из среды, то, очевидно, в ней установятся какие-то более высокие концентрации лимитирующих ресурсов. Точку, соответствующую концентрациям ресурсов в отсутствие потребления, Д. Тильман предложил называть точкой снабжения (англ. supply point). Фактически в неявном виде мы уже использовали это понятие, когда обсуждали модели, изображенные на рис. 58—59, и говорили о той или иной наблюдаемой в среде концентрации ресурсов. На рис. 61 в пространстве двух незаменимых ресурсов нанесена точка снабжения (ее координаты S1, S2) и изоклина нулевого прироста для одного вида. В каждой точке, находящейся на данной изоклине, рождаемость, по определению, равна смертности, но это не означает, что соотношение в потреблении двух ресурсов обязательно точно равно их соотношению при поступлении в среду. Из каждой точки мы можем провести вектор потребления С, показывающий то направление, в котором популяция стремится сдвинуть пороговую концентрацию, и вектор снабжения U, направленный к точке снабжения и показывающий то соотношение ресурсов, которое установилось бы в среде при некотором ослаблении его потребления данной популяцией. Вектор потребления и вектор снабжения могут быть направлены в строго противоположные стороны (под углом 180°): в этом случае соответствующая точка на изоклине будет называться точкой равновесия ресурсов (точка Е на рис. 61). В других точках изоклины вектор потребления и вектор снабжения могут находиться под углом, меньшим, чем 180°: такое соотношение ресурсов будет неравновесным.
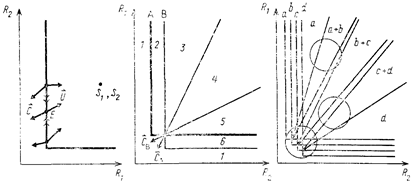
Рис. 61. Изоклина нулевого прироста популяции в координатах незаменимых ресурсов (по Tilman, 1982)
Рис. 62. Изоклины двух видов, ограниченных двумя незаменимыми ресурсами: CA и СB — векторы потребления (по Tilman, 1982)
Рис. 63. Изоклины четырех видов (а, b, с, d), ограниченных двумя ресурсами. Каждый из кружков показывает определенную вариабельность в количественном соотношении данных ресурсов в среде (по Tilman, 1982)
В случае пересечения изоклин двух видов, конкурирующих за два независимых ресурса, точка равновесия ресурсов — это как раз точка пересечения изоклин. На рис. 62 показаны векторы потребления (и продолжающие их векторы снабжения), исходящие из точки равновесия. Сосуществование видов в данном случае устойчиво, поскольку каждый из конкурирующих видов в большей степени потребляет тот ресурс, который сильнее ограничивает рост его собственной популяции. В частности, на рис. 62 вид А больше потребляет второй ресурс, а вид В — первый. Если бы ситуация была обратной, то сосуществование видов было бы неустойчивым. Если обратиться к схеме, изображенной на рис. 62, где цифрами обозначены отдельные области, ограниченные изоклинами и векторами, то в области 1 ни вид А, ни вид В существовать не могут, в области 2 А может существовать, а В не может; а области 6 наблюдается обратное положение — В может существовать, а А не может; в области 4 оба вида успешно сосуществуют; в области 3 А конкурентно вытесняет В, а в области 5 В конкурентно вытесняет А.
Вместо двух видов в пространстве двух ресурсов мы можем изобразить изоклины ряда видов и из точек пересечения этих изоклин провести векторы снабжения, ограничивающие области, в которых возможно сосуществование каждой пары видов (рис. 63). В разных точках этого пространства могут обитать один вид, два вида или ни одного. Иными словами, при точно определенном количественном соотношении двух ресурсов в каждом конкретном случае строго выполняется принцип конкурентного исключения: число сосуществующих видов не превышает числа лимитирующих ресурсов. Но если от идеализированной модели мы обратимся к природе, то обнаружим, что даже близко расположенные точки в любом реальном пространстве какого-либо местообитания (как наземного, так и водного) достаточно сильно различаются по количественному соотношению лимитирующих ресурсов. Кроме того, определенное для любой точки соотношение это может сильно меняться во времени. Так, например, проведенное Д. Тильманом очень подробное исследование распределения содержания азота в почве участка размером 12×12 м показало вариацию в 42 %, а вариация в содержании магния на том же участке достигала 100 %. Пространственно-временную вариабельность в поступлении ресурсов в среду на рис. 63 можно изобразить кружком определенного диаметра. Как видно из схемы, если этот кружок поместить в область высоких концентраций, то сосуществовать при таких вариациях могут не более двух видов, но если этот же кружок поместить в область низких значений, то он может покрыть область сосуществования сразу целого ряда видов. Иными словами, при очень низких концентрациях лимитирующих ресурсов даже весьма незначительной изменчивости их от одной точки пространства к другой или от одного момента времени к другому достаточно, чтобы обеспечить реальную возможность сосуществования сразу большого числа видов (во всяком случае, гораздо большего, чем число лимитирующих ресурсов). Из этого следует еще один любопытный вывод: при обогащении среды ресурсами мы вправе ожидать уменьшения видового разнообразия. Такое сокращение количества видов и усиление численного преобладания немногих видов действительно наблюдаются как в водной среде (явление эвтрофирования), так и в наземной (обеднение видового состава лугов при длительном их удобрении).
Заключение
В природе любая популяция вида организмов вступает в сеть взаимосвязей с популяциями других видов: Отношения типа хищник — жертва (или ресурс — потребитель) и конкурентные отношения — одни из наиболее важных в жизни любых организмов и в то же время одни из наиболее изученных. При возрастании численности жертв наблюдается как функциональная реакция хищника (т. е. увеличение числа жертв, потребляемых в единицу времени одной особью хищника), так и численная (т. е. увеличение численности популяции хищника). Благодаря способности хищников к функциональной и численной реакции пресс их на популяцию жертв выступает как фактор, зависящий от плотности и потому обладающий регуляторным воздействием.
Согласно теории, разработанной математиками, система взаимосвязанных популяций хищника и жертвы, скорее всего должна демонстрировать колебательный режим, но даже в лабораторных условиях получить устойчивые колебания хищник — жертва практически очень трудно. В тех же случаях, когда это удается, исследователи, как правило, ограничивают количество пищи для жертвы или же создают сложное гетерогенное местообитание, в котором жертва и хищник могут мигрировать, причем скорость расселения жертвы несколько больше скорости расселения хищника. В природных условиях мы обычно видим только следование численности хищника за колебаниями жертвы, определяемыми другими факторами, не связанными непосредственно с воздействием данного хищника.
Эволюция хищника и эволюция жертвы всегда теснейшим образом взаимосвязаны. Один из возможных в эволюции путей защиты жертвы от пресса хищников — увеличение рождаемости (компенсирующее соответствующее возрастание смертности от хищника). Другие возможные пути: это стратегия избегания встреч с хищником или стратегия выработки морфологических, физиологических и биохимических средств защиты от него. Обе эти стратегии, направленные на непосредственное снижение смертности от хищника, сопряжены для жертвы с определенными тратами, которые в конечном итоге выражаются в снижении рождаемости. Эволюция хищника направлена на повышение собственной рождаемости и (или) снижение смертности, что почти всегда связано с возрастанием эффективности использования жертв.
Конкурентные отношения между популяциями разных видов возникают тогда, когда они остро нуждаются в одном ресурсе, имеющемся в недостаточном количестве. Протекать конкуренция может по типу эксплуатации, т. е. простого использования дефицитного ресурса, или же по типу интерференции, при которой особи одного вида создают помехи особям другого в использовании общих ресурсов.
В экологии существует давняя традиция теоретического исследования конкуренции. Согласно математической модели Вольтерры—Лотки, позднее развитой и подтвержденной экспериментально Г. Ф. Гаузе, два вида, конкурирующие за один ресурс, как правило, не могут устойчиво сосуществовать в гомогенной среде, а исход конкуренции определяется соотношением интенсивности самоограничения каждой из популяций и их взаимоограничения. Это правило, известное также как закон Гаузе, или принцип конкурентного исключения, в результате всестороннего изучения теоретиками и экспериментаторами претерпело определенное развитие. В современной формулировке оно гласит, что число видов, неограниченно долго сосуществующих в постоянных условиях гомогенного местообитания, не может превышать числа плотностнозависимых факторов, лимитирующих развитие их популяций.
Закон Гаузе продолжает сохранять эвристическое значение для натуралистов, изучающих конкуренцию в природе. Прямые доказательства важности роли межвидовой конкуренции в природе получить неизмеримо труднее, чем в лаборатории. Поэтому, как правило, о значении конкуренции в качестве фактора, определяющего динамику и распределение природных популяций, судят по совокупности косвенных свидетельств.
В ряде случаев число сосуществующих видов, конкурирующих за общие, лимитирующие их развитие ресурсы, явно больше, чем число таких ресурсов (примером может быть сообщество планктонных водорослей или сообщество луговых растений), что противоречит закону Гаузе. Это противоречие снимается, однако, теорией, учитывающей пространственную и временную вариабельность в обеспеченности конкурирующих видов лимитирующими ресурсами.
[1] На русском языке слово «экология» впервые было упомянуто, видимо, в кратком конспективном изложении «Общей морфологии» Э. Геккеля — небольшой книжки, вышедшей в 1868 г. под редакцией И. И. Мечникова.
[2] Сейчас мы, правда, начинаем осознавать, что, возможно, и ни к чему стремиться развивать экологию и биологию вообще по образцу физики. Не исключено, что биология будущего окажется ближе к гуманитарным наукам. Во всяком случае, «приспособленность» — одно из центральных понятий в дарвинизме (а это пока единственная достаточно общая эколого-эволюционная теория) — относится к области семантической информации (Заренков, 1984).
[3] Самая богатая по числу видов группа организмов на земле — насекомые. Видов насекомых значительно больше, чем всех остальных видов животных и растений, вместе взятых. Общее число их остается неизвестным, так как большинство обитающих в тропиках видов насекомых еще не описаны. До недавнего времени считалось, что насекомых 3—5 млн. видов, но в последние годы появились данные (May, 1988), свидетельствующие о том, что эту цифру надо увеличить, возможно, на порядок, т. е. число видов насекомых на Земле не менее 30 млн. Основанием для этой переоценки послужили, в частности, результаты обследования крон тропических деревьев. Так, применив методику фумигации для изгнания насекомых из крон, удалось показать, что на 19 экз. одного вида тропических вечнозеленых деревьев Geuhea seemanni в Панаме одних только жуков обитало 1100 видов.
[4] Приведенная дефиниция, как указывает автор, представляет собой несколько измененное определение экологии, предложенное австралийским исследователем Г. Андревартой (Andrewartha. 1961), который в свою очередь исходил из идей, развиваемых еще в 20-х гг. Ч. Элтоном (1934; Elton, 1927).
[5] Подобная ситуация наблюдалась, впрочем, и в физике. Как отметил Вайскопф (1977), прогресс, достигнутый этой наукой в Новое время, связан с отказом от попыток установить сразу всю истину и объяснить целиком всю Вселенную. Вместо того чтобы ставить общие вопросы и получать частные ответы, ученые стали задавать более частные вопросы, но, как ни удивительно, получали на них более общие ответы.
[6] Заметим, что выражения «достаточно совершенен» или «достаточно приспособлен» вовсе не означает того, что вид этот приспособлен наилучшим образом, и дальше совершенствоваться ему некуда. Из сказанного не следует также, что каждый вид живет в природе в наиболее оптимальных условиях. Нередко случается, что из наиболее оптимальных (по абиотическим условиям) участков своего потенциального ареала вид вытесняется конкурентами или хищниками. Достаточно сослаться хотя бы на приведенный выше пример со зверобоем, находящимся под сильным прессом листогрыза Chrysolina.
[7] В англоязычной литературе, посвященной эволюционным аспектам экологии, очень часто цитируется английская поговорка «Jack of all trades is a master of none», которую на русский можно примерно перевести так: «Тот, кто берется делать любую работу, не делает хорошо ни одну из них».
[8] Специалисты-систематики замечают (Скворцов, 1988), что определенные названия таксономических категорий — это скорее имена собственные, а не нарицательные. Например, сказав «класс однодольных» или «класс пресмыкающихся», мы прежде всего представляем себе однодольные и пресмыкающихся, а не некий «класс вообще» — условную единицу систематиков, договорившихся, что классы делятся на отряды, а объединяются в типы.
[9] Среди отечественных ученых такую точку зрения отстаивал С. С. Шварц (1969). Близкого мнения придерживается в настоящее время А. В. Яблоков (1987), который в своей книге «Популяционная биология» определяет популяцию как «...минимальную самовоспроизводящуюся группу особей одного вида, на протяжении эволюционно длительного времени населяющую определенное пространство, образующую самостоятельную генетическую систему и формирующую собственное экологическое пространство» (с. 150). Поясняя свое определение, А. В. Яблоков подчеркивает, что «...популяция — это всегда достаточно многочисленная группа особей, на протяжении большого числа поколений в высокой степени изолированная от других аналогичных групп особей» (с. 151).
[10] Клонами обычно называют группы особей, произошедших от одной предковой формы путем вегетативного или партеногенетического размножения ч потому являющихся очень близкими родственниками. Экологи очень часто используют в своих опытах клоны водорослей, простейших, коловраток и других организмов.
[11] Подобной точки зрения часто особо строго придерживаются фитоценологи. Вместо термина «популяция» они предпочитают использовать термин «ценопопуляция», подчеркивая тем самым, что это не просто совокупность растений определенного вида, а совокупность, входящая в конкретный ценоз (= сообщество).
[12] Н. П. Наумов в 1960-е гг. последовательно отстаивал «мягкое» определение популяции, справедливо подчеркивая, что сами споры о возможности или невозможности считать ту или иную группировку популяцией имеют объективный характер, поскольку отражают естественную иерархическую структуру популяции. На наш взгляд, совершенно справедливо Н. П. Наумов (1965, с. 626) считал, что динамика численности — это «явление, разворачивающееся не только во времени, но и в пространстве».
[13] Оценка общей численности популяции особенно важна для исчезающих, занесенных в Красную книгу видов животных и растений. Вопрос о том, каким может быть минимальный допустимый размер этих популяций, приобретает уже сугубо практический характер.
[14] Специалисты, изучающие методику опенки пространственного распределения, рекомендуют применять показатель σ2/ т только в тех случаях, когда при увеличении среднего (что достигается использованием более крупных площадок) дисперсия растет по линейному закону. В других случаях используют иные показатели пространственной агрегированности (Романовский, 1979).
[15] Подчеркнем, что в этом примере имеется в виду сухая масса пищи (сырая масса может быть в 10 раз больше). Все цифры заимствованы из обобщающей работы Б. Д. Абатурова и В. Н. Лопатина (1987).
[16] Интересно, что паразитические черви Plagiorhynchus cylindraceus (из класса скребней) меняют поведение мокриц таким образом, что они не задерживаются в укрытиях, в результате чего гораздо чаще становятся жертвой насекомоядных птиц — окончательных хозяев этого паразита (Мур, 1984).
[17] Особенно много исследований, доказывающих, что в тех или иных конкретных случаях именно биотические факторы ограничивают распределение организмов, выполнено на протяжении последних 20—25 лет. Напомним, однако, сколь важную роль разного рода биотическим взаимодействиям в поддержании «экономии природы» придавал еще в прошлом веке Ч. Дарвин. Убеждение Дарвина основывалось главным образом на интуиции натуралиста, но, конечно, ему был известен (об этом есть прямое упоминание в «Происхождении видов...») и богатый опыт ботанических садов. Сама возможность существования элементов чужеродной флоры в открытом грунте (конечно, при надлежащем уходе и защите от животных-фитофагов и растений-конкурентов) и вместе с тем крайняя редкость проникновения этих элементов в окружающие естественные биоценозы уже являются свидетельствами важности биотических взаимодействий как фактора, ограничивающего распространение организмов. Позднее, уже в 50-х гг. нашего века, этот тезис развивал Ч. Элтон (1960), собравший очень большое число данных о случаях удачной и неудачной интродукции видов из одной географической области в другую. Оказалось, что при переносе с одного материка на другой (например, из Евразии в Америку или наоборот) большинство интродукций оканчивается неудачей, тогда как значительно чаще удачные интродукции наблюдаются при переносе организмов на острова (что в свою очередь объясняется ослаблением в островных сообществах пресса хищников, паразитов и конкурентов).
[18] Закон (или правило) минимума носит имя Ю. Либиха (1803—1873) — выдающегося немецкого химика, заложившего основы агрохимии и теории минерального питания растений. Заметим, что сам Либих ни о каком законе не говорил. Выражение «закон минимума» появилось позже. Основанием для его формулировки послужили, видимо, некоторые тезисы, опубликованные Либихом в 1855 г. Приведем их ниже в переводе Д. Н. Прянишникова (из его предисловия к книге Либиха): «Если в почве или в атмосфере один из элементов, участвующих в питании растений, находится в недостаточном количестве или не обладает достаточной усвояемостью, растение не развивается или развивается плохо. Элемент, полностью отсутствующий или не находящийся в нужном количестве, препятствует прочим питательным соединениям произвести их эффект пли, по крайней мере, уменьшает их питательное действие... Прибавляя к почве отсутствующий или не находящийся в должном количестве элемент или обеспечивая переход его из нерастворимого состояния в растворимое, восстанавливают эффективность других элементов. Отсутствие или недостаток одного из необходимых элементов при наличии в почве всех прочих делает последнюю бесплодной для всех растений, для жизни которых этот элемент необходим» (Либих, 1936, с. 19). Очевидно, что в предлагаемой формулировке закон минимума относится только к незаменимым ресурсам (точнее, элементам питания). В дальнейшем оно стало применяться и к заменимым ресурсам, а потом и вообще к любым экологическим факторам. Так, существует, например, следующее определение закона минимума: «Распространение вида контролируется тем фактором окружающей среды, по отношению к которому организм имеет наиболее узкую зону приспособляемости или
|
|
|
