 |


Три функциональных блока мозга
|
|
|
|
[…]
Можно выделить три основных функциональных блока, или три основных аппарата мозга, участие которых необходимо для осуществления любого вида психической деятельности. С некоторым приближением к истине их можно обозначить как:
1) блок, обеспечивающий регуляцию тонуса и бодрствования;
2) блок получения, переработки и хранения информации, поступающей из внешнего мира;
3) блок программирования, регуляции и контроля психической деятельности.
Каждый из этих основных блоков имеет иерархическое строение и состоит по крайней мере из надстроенных друг над другом корковых зон трех типов: первичных (или проекционных), куда поступают импульсы с периферии или откуда направляются импульсы на периферию, вторичных (или проекционно-ассоциативных), где происходит переработка получаемой информации или подготовка соответствующих программ, и, наконец, третичных (или зон перекрытия), которые являются наиболее поздно развивающимися аппаратами больших полушарий и которые у человека обеспечивают наиболее сложные формы психической деятельности, требующие совместного участия многих зон мозговой коры.
Рассмотрим строение и функциональные особенности каждого из этих блоков головного мозга в отдельности.
1 Блок регуляции тонуса и бодрствования
Для того чтобы обеспечивалось полноценное протекание психических процессов, человек должен находиться в состоянии бодрствования. Известно, что только в оптимальных условиях бодрствования человек может принимать и перерабатывать информацию, вызывать в памяти нужные избирательные системы связей, программировать свою деятельность и осуществлять контроль за протеканием своих психических процессов, корригируя ошибки и сохраняя направленность своей деятельности.
|
|
|
Хорошо известно, что в состоянии сна четкая регуляция психических процессов невозможна, всплывающие воспоминания и ассоциации приобретают неорганизованный характер, и направленное избирательное (селективное) выполнение психической деятельности становится невозможным.
О том, что для осуществления организованной, целенаправленной деятельности необходимо поддерживать оптимальный тонус коры, говорил еще И.П. Павлов, гипотетически утверждавший, что если бы мы могли видеть, как распространяется возбуждение по коре бодрствующего животного (или человека), мы наблюдали бы «светлое пятно», перемещающееся по коре мозга по мере перехода от одной деятельности к другой и олицетворяющее пункт оптимального возбуждения.
[…]
И.П. Павлов не только указал на необходимость оптимального состояния мозговой коры для осуществления организованной деятельности, но и открыл основные нейродинамические законы возникновения такого оптимального состояния. Как было показано многочисленными исследованиями павловской школы, процессы возбуждения и торможения, протекающие в бодрствующей коре, подчиняются закону силы, характеризуются определенной концентрированностью, уравновешенностью и подвижностью.
Эти основные законы нейродинамики неприложимы к состояниям сна или утомления. Это является результатом того, что в так называемых «тормозных», или «фазовых», состояниях тонус коры снижается и, как следствие, нарушается закон силы: слабые раздражители уравниваются с сильными по интенсивности вызываемых ими ответов («уравнительная фаза») или даже превосходят их, вызывая более интенсивные реакции, чем те, которые вызываются сильными раздражителями («парадоксальная фаза»), в отдельных случаях реакции сохраняются только в ответ на слабые стимулы, в то время как сильные раздражители вообще перестают вызывать какие-либо ответы («ультрапарадоксальная фаза»). Кроме того, по мере снижения тонуса коры нарушается нормальное соотношение возбудительных и тормозных процессов и та подвижность, которая необходима для протекания нормальной психической деятельности. Все это указывает на то, какое решающее значение имеет наличие оптимального тонуса коры для организованного протекания психической деятельности.
|
|
|
Возникает, однако, вопрос: какие аппараты мозга обеспечивают поддержание оптимального тонуса коры, о котором мы только что говорили? Какие участки мозга регулируют и изменяют тонус коры, сохраняя его на нужное время и повышая его, когда в этом возникает необходимость?
Одним из наиболее важных в этом плане открытий было установление того факта, что аппараты, обеспечивающие и регулирующие тонус коры, могут находиться не в самой коре, а в лежащих ниже стволовых и подкорковых отделах мозга и что эти аппараты находятся в двойных отношениях с корой, тонизируя ее и в то же время испытывая ее регулирующее влияние.
В 1949 г. два выдающихся исследователя — Мэгун и Моруцци — обнаружили, что в стволовых отделах головного мозга находится особое нервное образование, которое как по своему морфологическому строению, так и по своим функциональным свойствам приспособлено к тому, чтобы осуществлять роль механизма, регулирующего состояния мозговой коры, т. е. способно изменять ее тонус и обеспечивать ее бодрствование.
Это образование построено по типу нервной сети, в которую вкраплены тела нервных клеток, соединяющиеся друг с другом короткими отростками. По сети этого образования, названного ретикулярной формацией, возбуждение распространяется не отдельными, изолированными импульсами, не по закону «все или ничего», а градуально, постепенно меняя свой уровень и, таким образом, модулируя состояние всего нервного аппарата.
Одни из волокон ретикулярной формации направляются вверх, оканчиваясь в расположенных выше нервных образованиях — зрительном бугре, хвостатом теле, древней коре и, наконец, в тех образованиях новой коры, роль которых в организации сложной психической деятельности была отмечена в предыдущих главах. Эти образования были названы восходящей ретикулярной системой. Как обнаружили последующие наблюдения, она играет решающую роль в активации коры, в регуляции ее активности.
|
|
|
Другие волокна ретикулярной формации имеют обратное на правление: они начинаются от более высоко расположенных нервных образований — новой и древней коры, хвостатого тела и ядер зрительного бугра — и направляются к расположенным ниже структурам среднего мозга, гипоталамуса и мозгового ствола. Эти образования получили название нисходящей ретикулярной системы.
Они, как было установлено дальнейшими наблюдениями, ставят нижележащие образования под контроль тех программ, которые возникают в коре головного мозга и для выполнения которых требуется модификация и модуляция состояний бодрствования.
Оба раздела ретикулярной формации составляют единую вертикально расположенную функциональную систему, единый саморегулирующийся аппарат, построенный по принципу рефлекторного круга, который может обеспечивать изменение тонуса коры, но который вместе с тем сам находится под регулирующим влиянием тех изменений, которые наступают в коре головного мозга. Это аппарат пластичного приспособления к условиям среды в процессе активной деятельности.
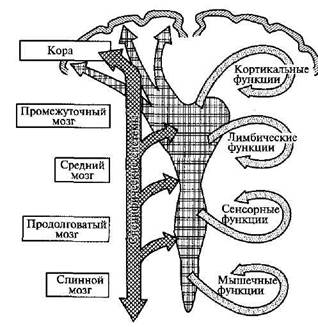
Рисунок: схема активирующей ретикулярной формации
С открытием ретикулярной формации в нейрофизиологию был введен фактически новый принцип — вертикальной организации всех структур мозга — и завершился длительный период, когда интерес ученых, пытавшихся найти нервные механизмы психических процессов, был сосредоточен лишь на аппаратах коры головного мозга, работа которой рассматривалась как не зависящая от нижележащих, глубоких образований. Ретикулярная формация — первый функциональный блок головного мозга — аппарат, обеспечивающий регуляцию тонуса коры и состояний бодрствования, аппарат, позволяющий регулировать эти состояния соответственно поставленным перед организмом задачам.
|
|
|
[…]
Активирующая ретикулярная формация, являющаяся важнейшей частью первого функционального блока мозга, с самого начала была названа неспецифической; это коренным образом отличало ее от других, специфических (сенсорных и двигательных) систем мозговой коры. Считалось, что ее активирующее и тормозное действие равномерно затрагивает как все сенсорные, так и все двигательные функции организма и что ее функцией является лишь регуляция состояний сна и бодрствования — того неспецифического фона, на котором протекают различные виды деятельности.
Это утверждение, которое было одним из исходных для классиков нейрофизиологии, впервые описавших активирующую систему мозга, нельзя, однако, признать полностью правильным. Как показали дальнейшие наблюдения, ретикулярная система мозга имеет определенные черты дифференцированности, или специфичности, как по своим анатомическим характеристикам (Бродал, 1957; М.Шейбел и А.Шейбел, 1958), так и по источникам активации и формам работы, однако эта специфичность не имеет ничего общего с «модальностью» органов чувств (анализаторов) (П. К.Анохин, 1958, 1961, 1963; Иошии, 1966, 1969).
Известно, что нервная система всегда находится в состоянии определенной активности и что наличие некоторого тонуса обязательно для любого проявления жизнедеятельности. Можно выделить по крайней мере два основных источника активации; действие каждого из них опосредствуется активирующей ретикулярной формацией, однако, что существенно, ее различными частя ми. В этом и состоит дифференцированность, или специфичность, функциональной организации «неспецифической активирующей системы». Первыми из источников активации являются обменные процессы организма, лежащие в основе гомеостаза (внутреннего равновесия организма) и инстинктивных процессов.
Обменные процессы (или, как иногда выражаются, «внутреннее хозяйство организма») в их наиболее простых формах связаны с дыхательными и пищеварительными процессами, с процессами сахарного и белкового обмена, с процессами внутренней секреции и т.д.; все они регулируются главным образом аппаратами гипоталамуса. Тесно связанные с гипоталамусом ретикулярные формации продолговатого мозга и среднего мозга играют значительную роль и этой наиболее простой, «витальной», форме активации.
Более сложные формы этого вида активации связаны с обменными процессами, которые организованы в определенные врожденные системы поведения; они широко известны как система инстинктивного (или безусловнорефлекторного) пищевого и полового поведения. Общим для обоих видов является то, что и в том и в другом случае источником активации являются обменные (гуморальные) процессы организма; отличаются они, во-первых, уровнем сложности организации и, во-вторых, тем, что если формы первого вида, наиболее элементарные, вызывают лишь примитивные автоматические реакции, связанные с недостатком кислорода или необходимостью выделения запасных веществ из их органических депо при голодании, то вторые предполагают организацию сложных поведенческих систем, в результате действия которых удовлетворяются определенные потребности и восстанавливается равновесие «внутреннего хозяйства организма».
|
|
|
Естественно, что для того чтобы обеспечить сложные инстинктивные формы поведения, необходима весьма избирательная специфическая активация, и биологически-специфические формы этой пищевой или половой активации являются функцией более высоко расположенных образований мезэнцефальной, диэнцефальной и лимбической систем. Большое число опытов, проведенных за последнее время, дают возможность убедиться в том, что в упомянутых образованиях мозгового ствола и древней коры имеются высокоспецифические ядра ретикулярной формации, раздражение которых приводит либо к активации, либо к блокированию различных форм инстинктивного поведения. […]
Второй источник активации имеет совсем иное происхождении. Он связан с поступлением в организм раздражений из внешнего мира и приводит к возникновению совершенно иных форм активации, проявляющихся в виде ориентировочного рефлекса.
Человек живет в мире информации, и потребность в ней иногда оказывается у него не меньшей, чем потребность в органическом обмене веществ.
Известно, что человек, лишенный обычного притока информации — что имеет место в редких случаях выключения всех воспринимающих органов, — впадает в сон, из которого его может вывести только поступление какой-либо информации. Известно, что нормальный человек тяжело переносит ограничение контактов с внешним миром, и, как это наблюдал Хэбб (1955), достаточно поместить испытуемых в условия резкого ограничения притока возбуждений (сенсорной депривации), чтобы у них возникли психические нарушения, галлюцинации, в какой-то мере компенсирующие это ограничение. В аппаратах ретикулярной формации имеются специальные механизмы, обеспечивающие тоническую форму активации, источником которой является главным образом приток возбуждений из органов чувств. Этот источник обладает не меньшей интенсивностью, чем первый, только что упомянутый, источник активации. Однако тоническая форма активации, связанная с работой различных органов чувств, является лишь наиболее элементарным источником активации того типа, который мы описываем.
Человек живет в условиях постоянно меняющейся среды, и это требует обостренного состояния бодрствования. Обострением бодрствования сопровождается всякое изменение в окружающих условиях, появление любого (как неожиданного, так и ожидаемого) события. Такая мобилизация организма лежит в основе особого вида активности, которую Павлов называл ориентировочным рефлексом и которая является основой познавательной деятельности.
Одним из наиболее важных открытий последних десятилетий было обнаружение связи ориентировочного рефлекса, или реакции пробуждения (активации), с работой ретикулярной формации мозга. Как показали исследования, ориентировочный рефлекс — сложное явление. Описаны тоническая и генерализованная формы реакции пробуждения, с одной стороны, и фазическая и локальная ее формы — с другой. Упомянутые формы ориентировочного рефлекса связаны с различными структурами в пределах ретикулярной формации: тоническая и генерализованная формы — с нижними, а фазическая и локальная — с верхними отделами ствола, и прежде всего с неспецифической таламической системой.
Как показали микроэлектродные исследования, неспецифические ядра зрительного бугра, а также хвостатого тела и гиппокампа функционально тесно связаны с системой ориентировочного рефлекса. Каждая реакция на «новизну» требует прежде всего сличения нового раздражителя с системой старых, уже появлявшихся ранее, раздражителей. Только та кое сравнение («компарация») может установить, является ли данный раздражитель действительно новым и требует ориентировочного рефлекса, или же уже знакомым, безразличным, не требующим специальной мобилизации организма. Только такой механизм может обеспечивать процесс «привыкания», когда многократно повторяющийся раздражитель теряет свою новизну и необходимость специальной мобилизации организма при его появлении исчезает. Очевидно, что в этом звене механизм ориентировочного рефлекса тесно связан с механизмами памяти, и именно связь обоих процессов обеспечивает компарацию сигналов, которая является одним из важнейших условий этого вида активации.
Важнейшим открытием последних лет было указание на тот факт, что значительная часть нейронов гиппокампа и хвостатого тела, не имеющих модально-специфических функций, осуществляют функцию компарации сигналов, реагируя на появление новых раздражителей и прекращая активность по мере привыкания к ним (О.С.Виноградова, 1969, 1970).
Активирующая и тормозящая, иначе говоря модулирующая, функция нейронов гиппокампа и хвостатого тела оказалась, как это стало ясным лишь в самые последние годы, основным источником регуляции тонических состояний мозговой коры, которые связаны с наиболее сложными формами ориентировочного рефлекса, носящими уже не врожденный, а прижизненно возникающий, или условнорефлекторный, характер.
Далее мы увидим, как влияет нарушение нормальной функции этих областей мозга на протекание психических процессов человека.
Остановимся в самых общих чертах на третьем и, пожалуй, наиболее интересном для нас источнике активации, в котором описанный только что функциональный блок мозга принимает самое непосредственное участие.
Источниками активности человека являются не только обменные процессы или непосредственный приток информации, вызывающий ориентировочный рефлекс. Значительная часть активности человека обусловлена намерениями и планами, перспективами и программами, которые формируются в процессе его сознательной жизни, являются социальными по своему заказу и осуществляются при ближайшем участии сначала внешней и потом его внутренней речи.
Всякий сформулированный в речи замысел преследует некоторую цель и вызывает целую программу действий, направленных на достижение этой цели. Достижение цели прекращает активность. Было бы, однако, неправильным считать возникновение намерений и формулировку целей чисто интеллектуальным актом. Осуществление замысла или достижение цели требует определенной энергии и может быть обеспечено лишь при наличии некоторого уровня активности.
Источники такой активности оставались долгое время неизвестными, и только исследования последних лет позволили сделать существенный шаг к тому, чтобы ответ на этот вопрос стал яснее.
Исследования, которые мы имеем в виду, заставляют отвергнуть старые предположения о том, что эти источники следует искать только во внутрикортикальных связях; они убедительно показывают, что в поиске механизмов наиболее высоких форм организации активности следует опираться на все тот же вертикальный принцип строения функциональных систем мозга и, следовательно, обратиться к связям между высшими отделами коры и нижележащей ретикулярной формацией.
До сих пор, обсуждая механизмы работы первого функционального блока, мы рассматривали только восходящие связи активирующей ретикулярной системы, однако мы упоминали, что наряду с ними существуют и нисходящие связи коры и нижележащих образовании; именно эти связи и осуществляют регулирующее влияние мозговой коры на нижележащие стволовые образования и являются механизмом, с помощью которого возникшие в коре функциональные узоры возбуждения вовлекают аппараты ретикулярной формации древнего мозга и получают энергетический заряд.
Нисходящие аппараты ретикулярной формации исследованы значительно меньше, чем восходящие связи. Однако благодаря целой серии работ выяснилось, что посредством кортико-ретикулярных путей раздражение отдельных участков коры может вызывать генерализованную реакцию пробуждения, оказывать облегчающее влияние на спинальные рефлексы, изменять — через систему гамма-афферентов — возбудимость мышц, повышать возбудимость кохлеарного аппарата, понижать пороги различительной чувствительности.
Таким образом, как морфологическими, так и морфофизио-логическими исследованиями с достаточной надежностью установлено, что наряду со специфическими сенсорными и двигательными функциями, на которых мы еще остановимся, кора головного мозга имеет и неспецифические активирующие функции и что раздражение определенных участков коры может оказывать как активирующие, так и тормозящие влияния на нижележащие нервные образования. Оказалось далее, что нисходящие волокна активирующей (и тормозящей) ретикулярной системы имеют достаточно Дифференцированную корковую организацию; если связанные со специфическими путями пучки волокон (повышающих или понижающих тонус сенсорных или двигательных аппаратов) исходят из первичных (и частично из вторичных) зон коры, то волокна, опосредствующие более общие активирующие влияния на ретикулярную формацию ствола, исходят прежде всего из лобных отделов коры. Нисходящие волокна, идущие преимущественно от префронтальной (орбитальной и медиальной) коры, адресуются к ядрам зрительного бугра и нижележащих стволовых образований и являются тем аппаратом, посредством которого высшие отделы мозговой коры, непосредственно участвующие в формировании намерений и планов, управляют работой нижележащих аппаратов ретикулярной формации таламуса и ствола, тем Самым модулируя их работу и обеспечивая наиболее сложные формы сознательной деятельности.
Все это показывает, во-первых, что аппараты первого функционального блока не только тонизируют кору, но и сами испытывают Н дифференцирующее влияние, и, во-вторых, что первый функциональный блок мозга работает в тесной связи с высшими отделами коры.
2 Блок приема, переработки и хранения информации
В предыдущем параграфе мы говорили об аппаратах первого функционального блока головного мозга, который играет важную роль в регуляции состояний активности коры и уровня бодрствования. Этот блок построен по типу неспецифической нервной сети, которая осуществляет свою функцию путем постепенного, градуального изменения состояний и не имеет непосредственного отношения ни к приему и переработке поступающей извне информации, ни к выработке намерений, планов и программ поведения. Этим первый функциональный блок мозга, расположенный в основном в пределах мозгового ствола, образований межуточного мозга и медиальных отделов новой коры, существенно отличается от аппаратов второго функционального блока мозга, основная функция которого заключается в приеме, переработке и хранении внешней информации.
Этот блок расположен в конвекситальных (наружных) отделах новой коры (неокортекса) и занимает ее задние отделы, включая в свой состав аппараты зрительной (затылочной), слуховой (височной) и общечувствительной (теменной) областей. По своему гистологическому строению он состоит из нейронов подкорки и мозговой коры. Эти нейроны в отличие от аппаратов первого блока работают не по принципу градуальных изменений, а по закону «все или ничего», принимая отдельные импульсы и передавая их на другие группы нейронов.
По своим функциональным особенностям аппараты этого блока приспособлены к приему экстероцептивных раздражений, при ходящих в головной мозг от периферических рецепторов, к дроблению их на огромное число компонентов (иначе говоря, к анализу их на мельчайшие составляющие детали) и к комбинированию их в нужные динамические функциональные структуры (иначе говоря, к синтезу их в целые функциональные системы).
Таким образом, этот функциональный блок головного мозга обладает высокой модальной специфичностью: входящие в его со став части приспособлены к тому, чтобы принимать зрительную, слуховую, вестибулярную или общечувствительную информацию в системы этого блока включаются и центральные аппараты вкусовой и обонятельной рецепции, но у человека они настолько оттесняются центральными представительствами высших экстероцептивных, дистантных анализаторов, что занимают в коре головного мозга незначительное место.
[…] основу этого блока составляют первичные, или проекционные, зоны коры, которые характеризуются высоким развитием нейронов IV афферентного слоя, значительная часть которых обладает высочайшей специфичностью. Ранее говорилось также, что нейроны зрительных аппаратов коры реагируют только на узкоспециальные свойствазрительных раздражителей (оттенки цвета, характер линий, на правление движения).
Эти высокодифференцированные нейроны строго сохраняю модальную специфичность, и в первичной затылочной коре можно найти лишь очень небольшое число клеток, которые реагируют на звук, прикосновение или вестибулярное раздражение.
Следует отметить, что первичные зоны отдельных областей коры, входящих в состав этого блока, включают в свой состав и некоторое количество клеток мультимодального характера, которые реагируют на несколько видов раздражителей и, по-видимому, выполняют функцию неспецифического поддержания тонуса; однако эти клетки составляют лишь небольшую часть всего нейронного состава первичных зон коры (по некоторым данным, количество их не превышает 4 – 5 %).
Над первичными, или проекционными, зонами коры разбираемого функционального блока мозга (составляющими его основу) надстроены, как мы видели ранее, аппараты вторичных, или гностических, зон коры, в которых IV афферентный слой уступает ведущее место II и III слоям, не имеющим столь выраженной модальной специфичности и включающим в свой состав значительное число ассоциативных нейронов с короткими аксонами, что позволяет комбинировать поступающие возбуждения в нужные «функциональные узоры» и осуществлять синтетическую функцию.
Подобное иерархическое строение, как уже отмечалось ранее, В той или иной степени свойственно всем областям коры, включенным во второй функциональный блок мозга.
В зрительной {затылочной) коре над первичными зрительными зонами (17-е поле Бродмана) надстроены вторичные зрительные Поля (18-е и 19-е поля Бродмана), которые, сохраняя свою модальную (зрительную) специфичность, работают в качестве аппарата, организующего те зрительные возбуждения, которые поступают в первичные зрительные поля.
С принципами работы зрительного анализатора мы познакомимся при рассмотрении нарушений зрительного восприятия, Наблюдаемых в клинике локальных поражений этих зон мозговой Коры.
Слуховая (височная) кора построена по тому же принципу. Ее первичные (проекционные) зоны скрыты в глубине височной коры и поперечных извилинах Гешля (41-е поле Бродмана). Нейроны этих зон высокодифференцированны и модально специфичны. Реагируют они только на те или иные звуковые раздражители. Как и первичное зрительное поле, первичные отделы слуховой коры имеют четкое топографическое строение. Как полагает ряд авторов, волокна, несущие возбуждения от тех отделов Кортиева органа, которые реагируют на высокие тона, располагаются во внутренних (медиальных), а волокна, реагирующие на низкие тона, — в наружных (латеральных) отделах Гешлевской извилины. Отличие первичных зон слуховой коры состоит в том, что соответствующие аппараты Кортиева органа представлены в проекционных зонах слуховой коры обоих полушарий, хотя преимущественно контралатералъный характер этого представительства все же сохраняется.
Над аппаратами первичной слуховой коры надстроены аппараты вторичной слуховой коры, которые расположены во внешних (конвекситальных) отделах височной области (22-, 42- и частично 21-е поля Бродмана) и которые также преимущественно состоят из II и III слоев клеток. Здесь, как и в соответствующих аппаратах зрительной коры, происходит превращение соматотопических проекций импульсов в их функциональную организацию. На детальном анализе функций этих зон и на картинах нарушений слухового восприятия, которые возникают при их поражении, мы остановимся далее.
Принципиально та же функциональная организация сохраняется, наконец, и в общечувствительной (теменной) коре. Как и в случае зрительной и слуховой коры, ее основу составляет первичная, или проекционная, зона (3-е поле Бродмана), где широко представлены высокомодально-специфичные нейроны IV слоя; для этого участка коры характерна и столь же четкая соматотопическая топография, в результате чего раздражение верхних участков этой зоны вызывает появление кожных ощущений в нижних конечностях, средних участков — в верхних конечностях контралатеральной стороны, а раздражение пунктов нижнего пояса этой зоны — ощущения в контралатеральных отделах лица, губ и языка.
Над первичной зоной общечувствительной (теменной) коры надстраиваются ее вторичные зоны (1-, 2- и 5-е поля Бродмана); так же, как и вторичные зоны зрительного и слухового анализаторов, эти зоны состоят преимущественно из нейронов II и III (ассоциативных) слоев, и их раздражение приводит к возникновению комплексных кожных и кинестетических ощущений. Эффект локального поражения их будет впоследствии предметом нашего специального рассмотрения.
Таким образом, основные, модально-специфические зоны разбираемого нами второго блока мозга, каждая из которых представляет собой центральный корковый аппарат того или иного модально-специфического анализатора, построены по единому принципу иерархической организации …
Как мы уже говорили ранее, познавательная деятельность человека никогда не протекает, опираясь лишь на одну изолированную модальность (зрение, слух, осязание); любое предметное восприятие (и тем более представление) является результатом поли модальной деятельности, которая первоначально в онтогенезе имеет развернутый характер и лишь затем становится свернутой. По этому естественно, что она должна опираться на совместную pаботу целой системы зон коры головного мозга.
Функцию обеспечения такой совместной работы целой группы анализаторов несут третичные зоны обсуждаемого нами блока, или, как их принято обозначать, зоны перекрытия корковых отделов различных анализаторов. Эти зоны расположены на границе затылочного, височного и заднецентрального отделов коры; их основную часть составляют образования нижнетеменной области, которая у человека развита настолько, что составляет едва ли не четвертую часть всех образований описываемого блока. Это дает основание считать третичные зоны специфически человеческими образованиями.
Как уже говорилось, третичные зоны задних отделов мозга состоят почти целиком из клеток II и III (ассоциативных) слоев коры и, следовательно, функция их почти полностью сводится к интеграции возбуждений, приходящих из разных анализаторов. Есть основания думать, что подавляющая часть нейронов этих зон имеет мультимодальный характер и реагирует на комплексные признаки среды (например, на признаки пространственного расположения, количество элементов), на которые не реагируют нейроны первичных и даже вторичных зон коры. Именно к таким третичным образованиям задних отделов мозговой коры относятся 7-, 39- и 40-е поля Бродмана (верхние и нижние отделы теменной области), 21-е поле височной и 37-е поле височно-затылочной областей.
Далее мы рассмотрим функции третичных образований описываемых отделов коры подробнее и убедимся на основании анализа психологических экспериментов и клинических данных, что в основном эти зоны связаны с пространственной организацией возбуждений и с превращением последовательно поступающих (сукцессивных) сигналов в одновременно действующие (симультанные) группы, чем и обеспечивается тот синтетический характер восприятия, о котором в свое время упоминал И.М. Сеченов (А.Р. Лурия, 1963, 1966).
Деятельность третичных зон задних отделов коры необходима не только для успешного синтеза наглядной информации, но и для перехода от уровня непосредственного наглядного синтеза к уровню символических процессов, для оперирования значениями слов, сложными грамматическими и логическими структурами, системами чисел и отвлеченными соотношениями. Другими словами, третичные зоны задних отделов коры являются аппаратами, участие которых необходимо для превращения наглядного восприятия в отвлеченное мышление, опосредствованное всегда внутренними схемами, и для сохранения в памяти организованного опыта.
Совокупность данных позволяет назвать этот функциональный блок мозга блоком получения, переработки и хранения экстероцептивной информации. Можно указать некоторые законы построения коры, входящей в состав второго блока мозга (этими законами описывается также и строение третьего блока).
Первый закон— закон иерархического строения корковых зон. Соотношение первичных, вторичных и третичных зон коры, осуществляющих все более сложный синтез приходящей информации, является достаточно отчетливой иллюстрацией этого закона.
Следует отметить, что отношения между первичными, вторичными и третичными зонами коры, входящими в состав этого блока, не остаются одинаковыми в процессе онтогенетического развития. Так, у маленького ребенка для успешного формирования вторичных зон необходима сохранность первичных зон, а для формирования третичных зон — достаточная сформированность вторичных зон коры. Поэтому нарушение в раннем возрасте низших зон коры соответствующих типов неизбежно приводит к недоразвитию более высоких зон коры; таким образом, как это было сформулировано Л.С. Выготским, основная линия взаимодействия этих зон коры направлена «снизу вверх».
Наоборот, у взрослого человека с полностью сложившимися психологическими функциями ведущее место переходит к высшим зонам коры. Даже воспринимая окружающий мир, взрослый человек организует свои впечатления в логические системы; иными словами, наиболее высокие, третичные, зоны коры у него управляют работой подчиненных им вторичных зон, а при поражении последних оказывают на их работу компенсирующее влияние. Такой характер взаимоотношений иерархически построенных зон коры в зрелом возрасте позволил Л.С. Выготскому заключить, что на позднем этапе онтогенеза они взаимодействуют «сверху вниз».
Второй закон можно сформулировать как закон убывающей специфичности иерархически построенных зон коры. Как уже было показано, первичные зоны коры каждой из частей, входящих в состав описываемого блока, обладают максимальной модальной специфичностью. Это характерно для первичных (проекционных) зон как зрительной, так и слуховой и общечувствительной коры. Основа этого явления — наличие огромного числа нейронов с высокодифференцированной, модально-специфической функцией.
Вторичные зоны коры, где преобладают верхние слои с их ассоциативными нейронами, обладают модальной специфичностью в значительно меньшей степени.
Будучи тесно связанными с корковыми отделами соответствующих анализаторов, эти зоны характеризуются модально-специфическими гностическими функциями. Здесь интегрируется в одних случаях зрительная (вторичные затылочные зоны), в других — слуховая (вторичные височные зоны), и третьих — тактильная информация (вторичные теменные зоны) Однако тот факт, что эти зоны (в которых, как уже говорилось, преобладают мультимодальные нейроны и нейроны с короткими аксонами) играют основную роль в превращении соматотипической проекции в функциональную организацию поступающей ин формации, указывает на то, что степень специализированности клеток этих зон значительно меньше, чем в первичных зонах, и что переход к ним знаменует существенный шаг в направлении «убывания модальной специфичности».
Еще в меньшей степени модальная специфичность характеризует третичные зоны описываемого блока, которые можно обозначить как «зоны перекрытия» корковых отделов различных анализаторов. Модальная специфичность входящих в их состав элементов представлена еще меньше, симультанные (пространственные) синтезы, которые осуществляются этими зонами коры, трудно расценить как модально-специфические. В еще меньшей степени этот принцип можно относить к высшим, символическим уровням работы этих зон, с переходом к которым функция третичных зон в известной мере приобретает надмодальный характер.
Таким образом, закон убывающей специфичности является другой стороной упомянутого нами ранее закона иерархического строения входящих в состав второго блока зон коры, обеспечивающих переход от дробного отражения частных модально-специфических признаков к синтетическому отражению более общих и отвлеченных схем воспринимаемого мира.
Следует отметить, что вторичные и третичные зоны коры (в которых пре
|
|
|
