 |


Целенаправленное действие и его компоненты 14 глава
|
|
|
|
Интересен вопрос о соотношениях общего количества генетической, поведенческой и логической информации. Однако что в этом случае принимать за количество генетической информации? Брать ли в качестве такого геном одного человека, или генофонд человеческой популяции, или сумму геномов всех существовавших когда-то и ныне живущих людей? Или, учитывая, что экологическая ниша человека включает в себя, в качестве субкомпонентов, экологические ниши всех видов организмов, понимать под общим количеством генетической информации совокупность генотипов всех, как вымерших, так и ныне здравствующих живых существ? Возможны разные варианты выбора, и все здесь зависит от точки зрения. Так, если сравнивать между собой только дискретные информационные системы, то количество кодирующей их генетической информации определяется лишь количеством ДНК в их геномах; количество информации поведенческой – информационной емкостью тех животных, которые такой информацией обладают; а количество логической информации – объемом всего человеческого знания, скомпонованного в максимально сжатый текст. Как соотносятся эти величины (выраженные в битах, напр.), сказать пока невозможно, но, по-видимому, третья во много раз превосходит обе предыдущие. Это и должно соответствовать все возрастающим размерностям пространства режимов, комплементарным информационным системам 1-го и 2-го рода.
В согласии с количеством кодирующей их информации, пространства режимов технологических операторов всегда и неизменно включали в себя пространства режимов меньших размерностей, комплементарные информации поведенческой и генетической. Говоря несколько иначе, это означает, что техногенез в своем развитии мог опираться только на предшествовавший биогенез, а техногенные операторы всегда и неизбежно включают в себя, в качестве компонентов, операторы биологического происхождения.
|
|
|
Действительно, начало развития технологий можно, например, датировать временем начала изготовления и использования каменных орудий труда. Но в «полный оператор» здесь, помимо человека, входили также и другие операторы генетической информации – клубни и плоды растений, мясо и шкуры животных, с помощью этих орудий добывавшиеся. Следующий араморфоз в эволюции техногенеза произошел с переходом отдельных человеческих сообществ к животноводству и земледелию. Главным компонентом таких техногенных операторов становились искусственно совершенствуемые сорта растений и породы животных, а ареной их деятельности, их экологической нишей оставались различные природные биоценозы. Эту линию рассмотрения можно было бы проводить и далее, но, пожалуй, достаточно отметить, что трансформируемые человеком фрагменты генетической информации до сих пор лежат в основе источников его питания техногенного происхождения, техническое обеспечение чего тесно переплетается с природными ландшафтами.
Но это лишь «одна сторона медали». Второй ее стороной являются побочные продукты w деятельности техногенных операторов. По мере роста численности человеческих сообществ и увеличения энергоемкости технологий побочные продукты их деятельности все сильнее деформируют природные популяции и биоценозы, вызывая в них неконтролируемые изменения, вплоть до угрожающих разрушением этих ценозов. Уже давно начала складываться парадоксальная ситуация: технологии, облегчающие человеку использование природных ресурсов, начали становиться главной причиной их разрушения. К настоящему времени этот процесс приобрел глобальный характер и именуется экологическим кризисом [38, 39].
|
|
|
Таковы, коротко, основные соотношения между техногенезом и биосферой.
Литература
1. Уоддингтон К. X. Основные биологические концепции. В кн.: На пути к теоретической биологии. 1. Пролегомены. М., «Мир», 1970.
2. Меллер Г. Ген как основа жизни. В кн.: Избр. работы по генетике, М.-Л., Огизсельхозгиз, 1937. С. 148-177.
3. Гершкович И. Генетика. М., «Наука», 1968.
4. Эйген М., Шустер П. Гиперцикл. М., «Мир», 1982.
5. Spiegelmann S. Quart. Rev. Biophys., 1971, V.4, p.213; Haruna I.,Spiegelmann S. Proc. Nat. Acad. ScL, USA, 1975, V.54, p.579.
6. Моисеев Н. Н. Расставание с простотой. М., «Аграф» (сер. Путь к очевидности), 1998.
7. Sumper M., Luse R. Proc. Nat. Acad. ScL, USA, 1975, V.72, p.1750.
8. Эйген М. Самоорганизация материи и эволюция биологических макромолекул. М., Мир, 1976.
9. Баблоянц А. Молекулы, динамика и жизнь. М., «Мир», 1990.
10. Кальвин М. Химическая эволюция. М., «Мир», 1971.
11. Печуркин Н. С. Энергия и жизнь. Новосибирск, «Наука», 1988.
12. Грант В. Эволюция организмов. М., «Мир», 1980.
13. Хакен Г. Синергетика. М., «Мир», 1980.
14. Кольцов Н. К. Журн. общ. биол., – 1972, т. 33, №4, С. 493.
15. Сарапульцев Б. И., Гераськин С. А. Генетические основы радиорезистентности и эволюции. М., Энергоатомиздат, 1993.
16. Иванов А. В. Происхождение многоклеточных животных. Л.,«Наука», 1968.
17. Маргелис Л. Роль симбиоза в эволюции клетки. М., «Мир», 1983.
18. Вернадский В. И. Биосфера. Статьи по биогеохимии. В кн.: Избр.соч., т. V, М., Изд. АН СССР, 1960.
19. Тейяр де Шарден П. Феномен человека. М., «Прогресс», 1965.
20. Дарвин Ч. Происхождение видов. В кн.: Собр. соч., т.l, M., Изд.Ю. Лепковского, 1907.
21. Серебровский А. С. Некоторые проблемы органической эволюции. М.,«Наука», 1973.
22. Тимофеев-Ресовский Н. В., Воронцов Н. Н., Яблоков А. Н. Краткий очерк теории эволюции. М., «Наука», 1969.
23. Корогодин В. И. Биофизика, 1983, т. 28, в. 1, С. 171-178.
24. Эрман Л., Парсонс П. Генетика поведения и эволюция. М., «Мир»,1984.
25. Морган Т. Г. Экспериментальные основы эволюции. М.-Л., Биомедгиз, 1936.
26. Кордюм В. А. Эволюция и биосфера. Киев, «Наумова думка», 1982.
27. Хесин Р. Б. Непостоянство генома. М., «Наука», 1984.
28. Поршнев Б. Ф. О начале человеческой истории (Проблемы палеопсихологии). М., «Наука», 1976.
29. Алексеев В. П. Становление человека. М., Изд. полит, лит., 1984.
30. Шовен Р. Жизнь и нравы насекомых. М., Гос. изд. селъхоз. лит.,1960.
|
|
|
31. Винер Н. Изобр. и рационал., 1974, №9, с.37.
32. Кропоткин П. А. Взаимная помощь среди животных и людей как двигатель прогресса. 2-е издание Пг.-М., 1922.
33. Дарвин Ч. О выражении душевных движений у человека и животных. В кн.: Собр. соч., т.Ш, М., Изд. Ю.Лепковского, 1908.
34. Швейцер А. Культура и этика. М., «Прогресс», 1973.
35. Моисеев Н. Н. Человек, среда, общество. М., «Наука», 1982.
36. Гессе Г. Игра в бисер. М., «Худ. лит.», 1969.
37. Уголев А. М. Естественные технологии биологических систем. Л.,«Наука», 1987.
38. Моисеев Н. Н. Алгоритмы развития. М., «Наука», 1987.
39. Моисеев Н. Н. Расставание с простотой. М.: «Аграф» (сер. Путь к очевидности), 1998.
Глава шестая
БИОСФЕРА И ТЕХНОСФЕРА
Биосфера и ее основные параметры
Как подчеркивал В. И. Вернадский [1], развитие человеческого мышления и все возрастающее его воздействие на окружающую среду – через порождаемые им технологии – следует рассматривать как природное явление, как неизбежное следствие цефализации. Эти процессы и привели к формированию техносферы – создаваемой самим человеком среды его обитания. Взаимоотношения биосферы и техносферы, в связи с грозящим Земле экологическим кризисом, являются сейчас объектом пристального внимания самых разных специалистов. Попробуем сформулировать на этот счет свою точку зрения, основанную на изложенных выше представлениях о природе информации и закономерностях динамики информационных систем.
Следуя традиции, биосферой будем называть всю населяющую Землю биоту (т.е. совокупность всех микроорганизмов, грибов, растений и животных) и среду ее обитания, включая почвенный покров и содержащие признаки жизни слои атмосферы. Накануне появления человека биосфера по своим основным параметрам вряд ли существенно отличалась от нынешнего ее состояния. Более того, такие ее характеристики, как общая масса живого вещества (порядка 2,4·1018 г); элементарный состав биомассы (азот – около 0,3%, углерод – около 3%, кислород – около 75%, водород – 10% и т.д.); содержание кислорода в атмосфере (около 21%, а всего порядка 1,2·1021 г); скорость круговорота слагающих ее атомов (для углерода, например, среднее время пребывания одной молекулы СО2 в атмосфере равно десяти годам); количество достигающей Земли солнечной энергии (167 ккал·см-2тод-1) – сохранялись неизменными, вероятно, на протяжении сотен миллионов лет [2-4]. Эта стабильность основных параметров биосферы обусловливается, с одной стороны, постоянством потока солнечной энергии, а с другой – развитой системой взаимоотношений между слагающими биосферу живыми компонентами, выполняющими определенные функции в «работе» по ее стабилизации (см., напр., [5]).
|
|
|
Косные и живые компоненты биосферы
Биосферу в целом можно подразделить на косные и живые компоненты. Косные компоненты – это химические соединения и физические тела, не входящие в данный момент времени в состав живых организмов. Это, прежде всего, газы, находящиеся в свободном состоянии (в атмосфере) или растворенные в водных бассейнах, вода в виде водяного пара, рек, озер, морей, океанов и ледников, различные неорганические и органические соединения, растворенные в этой воде и накапливающиеся в донных отложениях и почве, еще не претерпевшие полной деструкции отмершие компоненты живых организмов (листья, сучья, сброшенная при линьке шерсть и т.п.) и трупы самих этих организмов (от вирусов и бактерий до слонов и баобабов). По самой приблизительной оценке косные компоненты составляют более 99%, а на живые приходится меньше 1% от общей массы биосферы. Косные компоненты распределены по всей биосфере относительно диффузно. Значительное количество их включено в постоянный биологический круговорот, т.е. периодически входят в состав биоты. Химические соединения, выходящие из биологического круговорота и слагающие мощные осадочные породы, будем считать находящимися вне пределов биосферы.
В отличие от косных компонентов, живые компоненты биосферы четко структурированы. Элементарными структурами биосферы являются биоценозы, слагаемые, в свою очередь, трофически взаимодействующими популяциями, состоящими из отдельных индивидуумов.
По примерным данным, около 1% от всей биомассы Земли приходится на микроорганизмы и животных (2,3·1016 г), около 99% − на зеленые растения (2,4·1018 г). Только 0,13% биоты обитает в океанах, а остальные 99,87% − на континентах. В настоящее время на Земле известно около 100 тыс. видов микроорганизмов (бактерии, простейшие, грибы и водоросли), 150 тыс. видов высших растений и 1 млн. видов животных, из которых более 800 тыс. приходится на членистоногих. В отдельные биоценозы входят десятки и сотни взаимодействующих друг с другом видов, представленных популяциями, состоящими из сотен (для животных), тысяч (для растений) и миллионов (для микроорганизмов) особей. Жизнедеятельность этих последних (т.е. отдельных индивидуумов) и обусловливает, в конечном счете, стабильность характеристик биосферы и ее элементный состав.
|
|
|
Круговорот веществ в биосфере и факторы, его обусловливающие
Как мы уже отмечали, характерной особенностью элементного состава биосферы является его постоянный круговорот, т.е. переход из косного в живое и обратно. Можно полагать, что различные химические элементы, захваченные в период становления биосферы вихрем этого круговорота, лишь медленно вырываются из него, устремляясь в космическое пространство (молекулы газов) или выделяясь в виде водонерастворимых соединений, слагающих осадочные породы (известняки, сланцы), а также залежей углей, нефти и некоторых рудных месторождений. Столь же медленно им на смену в биосферу включаются новые атомы, извлекаемые хемотрофами из основных пород. Многократное «пропускание» через биологические компоненты одних и тех же атомов вещества есть один из основных законов функционирования биосферы. Но общее количество вещества, находящегося в «обороте», ограничено. Именно это, по-видимому, налагает основные ограничения на изменения количества живого вещества на нашей планете, определяя его постоянство.
Круговорот атомов в биосфере определяется тремя основными факторами: метаболизмом организмов, их размножением и их отмиранием. Во время становления биосферы, когда биомасса живого вещества возрастала, стремясь к своему пределу (на котором находится и по сей день), размножение, в общем, преобладало над гибелью и сопровождалось ростом дифференциации организмов по их трофическим функциям, т.е. увеличением числа видов. Этот период характеризовался вовлечением все большего количества атомов косного вещества в биологический круговорот. Однако по мере формирования биосферы, наряду с продолжающимися сменами ее видового состава и формированием все новых вариантов биоценозов, общая масса биоты возрастала все медленнее и наконец стабилизировалась на современном уровне. Произошло это не менее чем 2-3 млрд. лет тому назад.
Что положило предел этому процессу? Было ли это пределом возможности использования солнечной энергии или пределом доступности первичных источников атомов? Или это – гигантский аналог равновесия химических реакций, когда количество вновь синтезируемого продукта сравнивается с количеством разрушающегося? Или – результат насыщения «емкости» жизненного пространства? Как бы то ни было, можно полагать, что стабилизация количества биомассы на Земле была тесно связана с ускорением образования новых видов, т.е. с ускорением процесса эволюции.
Популяции
Эволюционирующими единицами являются не отдельные индивиды, а популяции [6], которые образуют информационные системы 1-го рода.
Как мы помним, каждый живой организм представляет собой информационную систему 1-го рода, состоящую из генетической информации и кодируемых ею операторов – его цитоплазматических и соматических компонентов. Популяции живых организмов составлены из множества информационных систем – организмов, связанных общностью происхождения и вписанных в тот или иной биоценоз, составляющий, вместе с окружающей средой, их экологическую нишу. Популяции организмов одного и того же вида могут быть включены не только в идентичные, но также в различающиеся биоценозы, в чем проявляется полипотентность присущей им генетической информации.
Каждую популяцию, из каких бы организмов она ни состояла, можно охарактеризовать двумя параметрами: средним временем τ1 удвоения числа слагающих ее особей в условиях данной экологической ниши s и средней продолжительностью их жизни τ2, в данных условиях. Параметр L = τ1/ τ2 = VP/Vr отражает надежность системы в условиях s, обусловленную особенностями организации операторов Q1 и, следовательно, задаваемую кодирующей их генетической информацией I. Если τ1> τ2, (т.е. L > I), то, благодаря размножению организмов, происходит возрастание численности слагаемой ими популяции; при τ1= τ2 (L = 1) наблюдается простое их воспроизведение, и численность популяции сохраняется на некотором постоянном уровне; в случае же τ1 < τ2 (L < I) преобладает отмирание, и численность популяции уменьшается.
Заметим, что при L = 1 состояние системы, представленной популяцией данных организмов Q1, становится нестабильным, точнее − губительным для этой популяции: любые флуктуации среды обитания, в том числе сдвиги внутри экосистемы, вызывающие даже временное превышение τ2, по сравнению с τ1, будут приводить систему к гибели.
Существует давно сформулированная теорема [7], согласно которой при L = 1 популяция обречена на гибель по чисто внутренним причинам. С течением времени численность такой популяции все более флуктуирует, и, когда она приблизится к нулю, популяция исчезнет. Конечно, время гибели популяции при L = 1 сильно зависит от ее исходной численности, но результат остается неизменным: даже при сколь угодно большой исходной численности п0 популяции при t → ∞ вероятность ее выживания р → 0. Конечно, уменьшение τ2, ускорит наступление гибели популяции. Поэтому можно утверждать, что стабильность или надежность популяции определяется неравенством L > 1 и тем выше, чем больше величина параметра L [8].
Давление жизни и жизнеспособность популяций
Утверждение, что при L = 1 информационная система обречена на гибель, очень сильное и поэтому требует специального обоснования. Условие L > 1 означает, что численность объектов, представляющих данную информационную систему, возрастает в геометрической прогрессии. В действительности, как мы знаем, этого не происходит – численность природных популяций в среднем стабильна. Значит, отношение L > 1 равносильно «давлению жизни» и отражает ту «силу», с которой данная популяция как бы противостоит разрушительному воздействию на нее различных внешних «помех».
Если численность популяции, по каким-либо причинам, снизилась ниже некоторого критического уровня, такая популяция, как правило, деградирует и исчезает. Только условие L > 1 позволяет популяции сохранять свою численность выше критического уровня при усилении давления извне (усилении помех) и достигать оптимального значения при ослаблении такого давления. В случае же L = 1 ослабление давления среды не будет сопровождаться ростом численности популяции, а любое усиление помех будет приводить к уже некомпенсируемому уменьшению ее численности и, в конце концов, к ее вырождению.
Экосистемы, их компоненты и характеристики
Чтобы яснее представить себе положение популяций в экосистемах, вспомним одно из основных свойств любого целенаправленного действия − сопровождающее его производство побочных продуктов w.
Компоненты, слагающие побочные продукты, по определению не могут быть реутилизированы образующими их в процессе своей жизнедеятельности организмами. Если w «обезвреживаются» в s, они не будут угрожать существованию своих «продуцентов». Но если w накапливаются в s, это неизбежно приведет к «отравлению» среды обитания таких продуцентов и, как следствие, к гибели их популяции, даже при избыточном содержании в этой среде ресурсов, необходимых для их самовоспроизведения: ни один организм не может существовать в среде, перегруженной отходами его жизнедеятельности [9].
Таким образом, условиями существования самовоспроизводящихся информационных систем являются: неуменьшение ресурсов R данной среды обитания s ниже некоторого предела, и неувеличение содержания в ней побочных продуктов w выше некоторого порогового значения.
Таким образом, w представляет собой биогенный компонент среды обитания организмов, угрожающий существованию тем из них, которые его производят.
Но это – лишь один аспект w. Второй его аспект − это то, что w, в принципе, могут быть утилизированы другими организмами, кодируемыми информацией с большим значением В и способными разрабатывать экологическую нишу более высокого яруса жизни (см. главу 5). Если бы этого не происходило, то, сколь бы велико ни было жизненное пространство s, со временем оно оказалось бы перегружено продуктами жизнедеятельности организмов исходной популяции и вместо «кишащего жизнью бассейна» превратилось бы в «выгребную яму».
Однако возникновение (или миграция из других местообитаний) в зоне 5 организмов, способных утилизировать w, сопутствующие жизнедеятельности ее аборигенов, будет приводить к формированию сообщества из двух популяций организмов. Альянс окрепнет, если продукты жизнедеятельности новичков окажутся в какой-либо мере «полезными» для аборигенов. Так сформируется простейшая экосистема, численность обитателей которой (или, точнее, плотность заселения) будет регулироваться обратными связями. Очевидно, что число живых компонентов экосистемы, т.е. число слагающих ее популяций организмов разных видов, может быть сколь угодно велико, но в реальных ситуациях будет определяться ее емкостью, запасами ресурсов R и трофическими взаимодействиями ее обитателей. Реальные экосистемы или биоценозы могут содержать сотни компонентов [9].
Таким образом, важнейших характеристик экосистемы две: число слагающих такие экосистемы видов N и средняя численность особей в представляющих эти виды популяциях ni (i = 1, 2,..., N).
Направления эволюции экосистем
Любую экосистему можно определить как систему самовоспроизводящихся популяций, занимающих единый ареал обитания и выполняющих две основные функции. Первая – обеспечение прироста численности популяций («давление жизни»), или общей биомассы V экосистемы (продуктивность данной экосистемы):  . Вторая функция – кондиционирование, или очистка среды обитания от побочных продуктов
. Вторая функция – кондиционирование, или очистка среды обитания от побочных продуктов 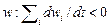 (i = 1, 2,..., N). Очевидно, чем больше скорость утилизации отходов, тем лучше условия для прироста биомассы, т.е. тем сильнее будет давление жизни.
(i = 1, 2,..., N). Очевидно, чем больше скорость утилизации отходов, тем лучше условия для прироста биомассы, т.е. тем сильнее будет давление жизни.
Условие равновесного существования экосистемы можно сформулировать как
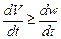 (22)
(22)
Оптимальным, по крайней мере для замкнутых экосистем с постоянным местообитанием, будет условие, когда кондиционирующая мощность экосистемы превосходит ее продуктивность, что соответствует поддержанию экосистемы в состоянии «максимальной чистоты».
Неравенство (22) представляет собою условие, определяющее направление эволюции популяций, слагающих любые экосистемы.
Это условие может реализоваться двумя путями: путем расширения ареала обитания данной экосистемы (экстенсивный путь ее развития) и путем увеличения ее кондиционирующей мощности (интенсивный путь). Третий возможный путь усиления неравенства (22) – уменьшение продуктивности экосистемы – нельзя считать приемлемым: уменьшение продуктивности может осуществляться лишь ценой понижения давления жизни слагающих экосистему популяций, что, как мы видели, чревато их деградацией и гибелью.
Так как в реальных ситуациях ареал обитания любой экосистемы всегда ограничен, то с течением времени на смену ее экстенсивный развитию, или экспансии, неизбежно должен прийти интенсивному путь, реализующийся через все большую «пришлифовку» друг к другу слагающих экосистему популяций. Это, кстати, и реальный путь эволюции видов, «эволюционирующими единицами» которых, как мы уже упоминали, являются именно популяции. Этот путь развития можно характеризовать изменениями числа и численности слагающих экосистему популяций.
Надежность экосистем, биоценозов и биосферы
Для любой экосистемы, представляющей собой совокупность N популяций разных видов, будет справедливо соотношение
 (23)
(23)
где V – биомасса данной экосистемы, G – константа скорости прироста биомассы, g - скорость потери биомассы, а φ и ψ - параметры, отражающие конкуренцию в этой системе. Это соотношение, по существу, является условием надежности экосистемы. Несколько (N) экосистем, связанных общим местообитанием, образуют один биоценоз. Для процветающего биоценоза справедливо неравенство
 (24)
(24)
Биоценоз как самоорганизующаяся система, состоящая из нескольких экосистем, может достигнуть стабильности и замкнутости при условии
 (25)
(25)
когда скорость прироста биомассы в каждой экосистеме и популяции равна скорости образования побочных продуктов в режиме их реутилизации.
Заметим, что условие стабильности биоценозов, в отличие от условия стабильности популяций, когда τ1 = τ2 не является для биоценозов пагубным, а напротив, это единственный залог их нормального функционирования. Это же справедливо и для всей биосферы, представляющей собой саморегулирующуюся и самостабилизирующуюся совокупность всех населяющих Землю биоценозов. При этом должно соблюдаться соотношение
 (26)
(26)
где п - численность каждой из N популяций (i = 1,2,... N), слагающих биосферу; Gi, и Di – скорости размножения и гибели особей, входящих в эти популяции; U i – параметр конкуренции особей i -ой популяции между собой; ψi,j - представленный в виде матрицы параметр, характеризующий взаимную утилизацию отходов организмами i -ой и j -ой популяций.
Соотношение (26) – условие стабильного функционирования совокупности экосистем, состоящих из большого числа видов, в том числе и всей биосферы. Очевидно, что надежность Ω этого функционирования обеспечивается надежностью выполнения как функций продуктивности (θ), так и функций кондиционирования (ø). Поэтому общая надежность может быть определена как произведение надежностей выполнения обеих функций, т.е. как Ω = θ·ø-ф.
Из сказанного выше следует основное требование к особенностям деятельности «функциональных аналогов», т.е. популяций разных видов, выполняющих в биоценозе или биосфере сходные экологические функции: функциональные аналоги могут заменять друг друга в данной экосистеме лишь при условии ΔΩ ≤_0. В противном случае (т.е. если ΔΩ < 0) такая замена окажется неполноценной и будет приводить либо к деградации, либо к перестройке любого биоценоза, а в общем случае – и всей биосферы.
Популяции в экосистемах: ограничения их численности и направления эволюции
Вернемся теперь к отдельным популяциям, составляющим экосистемы. Заметим, прежде всего, что экосистемы, как и экологические ниши, для разных организмов чаще всего перекрываются, но не полностью совпадают. Для биосферы в целом характерна иерархическая структура, так что обитатели верхних ярусов жизни, как правило, венчают собой или объединяют по нескольку экосистем, представленных обитателями низлежащих ярусов.
Законы, управляющие динамикой популяций разных организмов, едины, независимо от того, какую функцию они выполняют в биосфере и на каком уровне организации (и ярусе жизни) они находятся, т.е. будь это хемотрофы, ауксотрофы или гетеротрофы, продуценты или консументы, микроорганизмы, высшие растения, беспозвоночные или млекопитающие. Обусловлено это тем, что, независимо от видовой принадлежности, ни один организм не в состоянии выполнять обе основные функции в своей экосистеме – и функцию производства биомассы, и функцию утилизации побочных продуктов своей жизнедеятельности.
Обозначив численность популяции через п, напишем следующее уравнение:
 (27)
(27)
где G –коэффициент размножения данных организмов, D – средняя скорость их отмирания, a U –параметр конкуренции их друг с другом за среду обитания, питание и т.п. Показатель степени ψ при п отражает форму зависимости показателя конкуренции U от п, а член Unψ есть обобщенное выражение для обратной связи в популяции. Подчеркнем, что уравнение (27) не относится к человеку, ставшему на техногенный путь развития, ибо в этом случае параметр конкуренции может иметь совершенно иную связь с численностью популяции п, нежели в случаях других обитателей биосферы.
Следует отметить, что величина ψ может быть как больше, так и меньше нуля. Если в первом случае (ψ > 0) это отражает возрастание внутривидовой конкуренции с увеличением п (например, борьбу за территорию), то во втором случае (ψ < 0) это будет отражать существование взаимопомощи между членами данной популяции. Поэтому особый интерес представляет зависимость ψ от п для разных видов.
Выражение (27), связывающее через D, U и ψ динамику размножения (или роста численности) популяции с состоянием (или статусом) включающей ее экосистемы, означает, что в любой реальной ситуации – в реальном биоценозе или биосфере в целом – рост численности популяций всегда и неизбежно ограничен: при (G-D) п = Unψ прирост популяции dn/dt = 0.
Выше мы видели, что условие dn/dt =0 есть условие нестабильности состояния популяции. В случае сохранения постоянства других параметров экосистемы популяция при dn/dt = 0 была бы обречена на вымирание. Однако, благодаря наличию в любой экосистеме обратных связей, уменьшение численности данной популяции (если оно не ниже критического) неизбежно будет приводить к такому изменению статуса экосистемы, при котором условия обитания этой популяции опять сдвинутся в «благоприятную» сторону, что приведет к увеличению ее численности, за чем последует ее уменьшение, затем – опять увеличение и т.д. В реальной ситуации это будет проявляться как «волны жизни» [10], простейшей моделью которых может служить система «хищник-жертва» [11]. Поэтому в любой стабильной экосистеме численность любой популяции будет всегда колебаться около некоторого константного значения, задаваемого сформулированным выше правилом (27), не выходя за определенные максимальные и минимальные пределы.
|
|
|
