 |


Поперечнополосатая мускулатура
|
|
|
|
ВВЕДЕНИЕ
Физиология — наука о жизнедеятельности целостного организма и отдельных его частей: клеток, органов и систем. Физиология изучает работу органов дыхания, пищеварения, кровообращения, органов, осуществляющих осморегуляцию, обеспечивающих иммунитет, приспособление к меняющимся условиям обитания; изучает и суммирует сведения о количественном выражении обмена веществ, баланса энергии, действии нервных и гормональных механизмов управления жизнедеятельностью.
Существование физиологии рыб как отдельной науки обусловлено своеобразием условий обитания и большим хозяйственным значением рыб.
Многообразие экологии рыб:
| Размер | От нескольких метров до 2 мм |
| Температура | От температур, близких к точке замерзания до 40°С. |
| Питание | Хищники, растительноядные, детритофаги, планктофаги |
| Дыхание | Дышащие атмосферным воздухом, и виды, извлекающие кислород из воды, некоторые виды рыб способны значительное время существовать в среде, совершенно не содержащей кислорода |
| Соленость | Населяют и пресные воды, и водоемы с соленостью, превышающей океаническую |
| Размножение | Живородящие виды, откладывающие икру; некоторые популяции рыб, размножаются гиногенетически, т.е. без участия самцов |
| Индивидуальное развитие | Во время эмбрионального и личиночного периодов жизни рыб многие функции, такие, как дыхание, питание, осморегуляция, осуществляются совершенно иначе, чем во взрослом состоянии. Дыхание эмбрионов и личинок происходит всей поверхностью тела, а у взрослых рыб имеются жабры и даже легкие. Эмбрионы питаются желтком, запас питательных веществ, в котором во много раз больше, чем в самом эмбрионе. Взрослые же рыбы фильтруют, заглатывают, откусывают, жуют, сосут и другими способами поглощают и усваивают самую разнообразную пищу. |
|
|
|
Физиологию рыб невозможно изучать на каком-то одном объекте, настолько мир рыб разнообразен и удивителен.
Рыбоводство использует данные физиологии для рационализации кормления, составления полноценных рационов, удешевления кормов. На физиологических знаниях основаны мероприятия по стимуляции созревания половых продуктов рыб. Экологические расчеты и построения основываются на сведениях о питании, усвоении пищи, балансе вещества и потребностях разных видов рыб.
Нередко требуются физиологические данные для сравнения между собой различных видов рыб по использованию питательных веществ, по характеру нервных реакций, по скорости плавания, скорости изменения плавучести и т. д.
Основными методическими приемами в физиологии рыб являются измерение и эксперимент. Измеряется частота периодических процессов — дыхательных актов, сердце биения, желудочной моторики, плавательных движений. Определяются количественные показатели физиологических процессов — количество поедаемой пищи, количество прогоняемой через жабры воды, интенсивность кровотока, мочеобразования, секреции желудочного сока и т.д. Однако при измерениях необходимы особые приемы, учитывающие водный образ жизни рыб.

Изучаемых рыб содержат в аквариумах. Нормальную рыбу для физиологических исследований можно получить, если в аквариуме ей обеспечиваются нормальные условия существования. От культуры аквариальньгх работ в значительной степени зависит успех исследований.
Многие наблюдения и измерения проводят сквозь стенку аквариума, через поверхность воды. Визуальные наблюдения позволяют определять частоту дыхательных движений, частоту работы плавников, скорость перемещения рыб, частоту различных поведенческих актов — питания, стычек, ухаживания и т.д. Определяется длительность времени пребывания в различных зонах аквариума — на свету и в темноте, в условиях температурного градиента, в струях, содержащих пахучие и токсические вещества. Так изучают спектральную, звуковую, вкусовую чувствительность, а также характеристики образования навыков и рефлексов.
|
|
|
При проведении хирургических экспериментов у рыб вскрывают брюшную полость, обнажают мозг, нервы, экстирпируют железы внутренней секреции, делают кишечные и желудочные фистулы, канюлируют мочеточники и кровеносные сосуды. У рыб нет крупных подкожных кровеносных сосудов, как у людей и таких животных, как крысы, кролики и другие млекопитающие. Поэтому даже взятие крови у рыб требует специальных приемов.
Кровь у рыб берут из хвостовой вены, из сердца или из жаберной, артерии, используя пастеровские пипетки и шприцевые иглы. Гораздо сложнее, чем млекопитающим, вводить рыбам внутривенно различные вещества. Поэтому инъекции рыбам обычно производят внутримышечно, внутрибрюшинно или через рот. Можно вводить изучаемые вещества рыбам ректально. Для улучшенияисхода операций рыб промывают и содержат в растворах антибиотиков.
Определенные сложности возникают при изучении питания, пищеварения и экскреции у рыб. Корма, задаваемые рыбам, быстро размокают и теряют свои питательные свойства, так как многие вещества хорошо растворимы в воде. Это касается в первую очередь искусстве иных кормов — фаршей, порошков, гранул.
Поэтому для определения точного количества съеденной пищи используют вскрытие рыб, промывание пищеварительного тракта через рот или анус, принудительное кормление кусками пищи (применяют при изучении пищеварения хищных рыб), а также введение пищи с помощью зондов и трубок (используют при работе с мирными рыбами, имеющими узкий пищевод).
Определение усвоения рыбами питательных веществ корма связано со сбором фекалий, что очень трудно в воде. Поэтому прибегают к выдавливанию и отсасыванию содержимого заднего отдела кишечника, вскрытию рыб, или используют аквариумы с коническим дном и съемным сборником фекалии. Для определения усвоения корма в него вводят инертные (неусвояемые) вещества — целлюлозу, мелкие хорошо заметные частицы, окись хрома и другие вещества.
|
|
|
Изучение дыхания рыб отличается от изучения такового у животных, дышащих кислородом воздуха. Рыб помещают в замкнутые или проточные сосуды и определяют убыль растворенного в воде кислорода. Распространение в последние годы кислородных электродов сделало изучение дыхания рыб простым и удобным. Изучено потребление кислорода различными видами рыб в покое и при движении, при голодании и в процессе питания. Современные установки позволяют постоянно регистрировать потребление кислорода рыбами в течение многих суток подряд, используя автоматическую запись результатов.
Мышечную деятельность обычно изучают в поведенческих экспериментах на свободно плавающей рыбе или на установках для изучения почти целой (например, обезглавленной) или целой рыбы. Локомоторные движения рыб изучают путем киносъемки в естественных условиях и в условиях экспериментальных бассейнов и различных гидродинамических устройств.
Физиологические исследования проводят не только с отдельными рыбами на уровне поведения организма в целом, но и с отдельными частями организма на тканевом или даже на клеточном уровне. Например, многие вопросы, связанные с пищеварением, были выяснены с использованием препаратов пищеварительного тракта — отрезков и лоскутов кишечника, желудка.
При изучении нервной деятельности рыб используют поведенческие методы и приемы электрофизиологии — запись электрических потенциалов отдельных нервных волокон, нервов и нервных тканей. Современная аппаратура позволяет регистрировать электроэнцефалограммы, кардиограммы и миограммы рыб в покое и движении.
Эксперимент является одним из самых действенных приемов физиологии рыб. Он заключается в создании для животного или отдельного органа, ткани, клетки различных условий и в определении влияния этих условий на результирующий показатель. Рыб помещают в различные температурные условия, в воду с различной соленостью, насыщением кислородом, содержанием токсикантов или биологически активных веществ.
|
|
|
Экспериментальные воздействия могут заключаться также в скармливании различных по составу и величине рационов, в инъекциях медикаментами, хирургических вмешательствах. В настоящее время все чаще используются машинная обработка данных, полученных экспериментальным путем, и математическое моделирование физиологических процессов.
ТЕМА 1: Движение
Механическое движение занимает важное место в жизненных процессах. Рыба перемещается в воде — плавает, прокачивает воду сквозь жаберный аппарат, сердце проталкивает кровь по сосудам, кишечник проталкивает пищу. Имеются и другие, менее заметные, но тоже очень важные формы механического движения — изменяется просвет кровеносных сосудов и зрачка глаз, сокращаются и растягиваются стенки желчного, пузыря, сжимаются и расслабляются кольцевые сфинктеры желудка и кишечника.
Форма тела
Форма тела должна обеспечивать рыбе возможность передвигаться в воде (среде значительно более плотной, чем воздух) с наименьшей затратой энергии и со скоростью, соответствующей ее жизненным потребностям.
Форма тела, отвечающая этим требованиям, выработалась у рыб в результате эволюции: гладкое, без выступов, тело, покрытое слизью, облегчает движение; шеи нет; заостренная голова с прижатыми жаберными крышками и сжатыми челюстями рассекает воду; система плавников определяет движение в нужном направлении. В соответствии с образом жизни выделено до 12 различных типов формы тела. Наиболее характерные представлены на рис. 2.
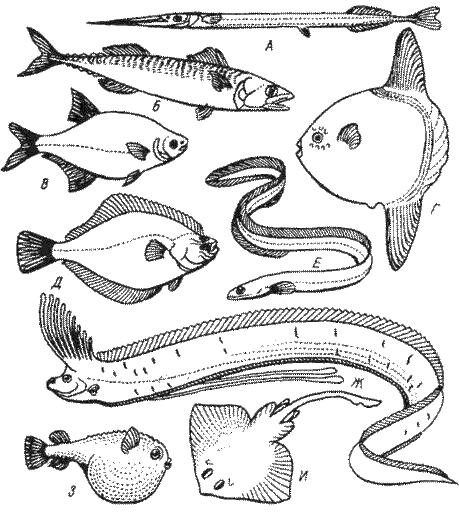
Рис. 2. Типы формы тела рыб.
А — стреловидный (сарган); Б — торпедовидный (скумбрия); В — сплющенный с боков, лещевидный (лещ обыкновенный); Г — тип рыбы луны (луна-рыба); Д — тип камбалы (речная камбала), Е — змеевидный (угорь); Ж — лентовидный (сельдяной король); 3 — шаровидный (кузовок); И — плоский (скат) (по Никольскому, 1974)
1. Торпедовидный (веретенообразный). Тело рыб похоже на торпеду или веретено, оно хорошо обтекаемо, немного сжато с боков и утончается к хвосту. Рыбы приспособлены к быстрому длительному плаванию в толще воды. Это наилучшие пловцы, совершающие продолжительные миграции к местам нагула и к местам икрометания (нерестилищам): тунец, макрель, сельдь, треска, лососи.
Такая же форма тела выработалась и у других хорошо плавающих водных животных, далеко отстоящих от рыб (китообразные и вымершие ихтиозавры).
2. Змеевидный. Тело вытянутое, змеевидное, округлое, на поперечном разрезе образует овал. Плавают, змеевидно изгибаясь всем телом. Это миноги, угри.
3. Лентовидный. Тело, подобное ленте, вытянуто вдоль, плоское с боков. Пловцы плохие, живут в спокойных водах больших глубин (сельдяной король, рыба-сабля).
|
|
|
4. Стреловидный. Тело удлиненное, сжато с боков, примерно одинаковой высоты; хвост сильный, голова заострена, спинной плавник сдвинут сильно назад. Эти рыбы продолжительных плаваний не совершают, но на небольшом расстоянии развивают огромную скорость, набрасываясь на добычу. Это хищники — щука, таймень.
5. Сплющенный. Здесь различают: а) симметрично-сжатую, лещевидную форму: тело высокое, сжатое с боков (лещ); б) несимметрично-сжатую: высокое, сжатое с боков тело несимметрично, глаза расположены на одной стороне (камбалы).
Такая форма тела не способствует быстрому перемещению, эти рыбы плохие пловцы.
6. Плоский. Тело сплющено в дорсовентральном направлении (сверху вниз). Обычно двигаются мало, живут у дна (скаты).
7. Шаровидный. Тело в виде шара, иногда передний отдел окружен костным панцирем (кузовок). Самостоятельно передвигаться иногда почти не могут. Это скалозубы — Tetrodon, Diodon.
Этими типами не исчерпывается многообразие форм тела рыб; у некоторых рыб форма тела является как бы промежуточной комбинацией нескольких типов.
Туловищный и хвостовой отделы тела рыб снабжены плавниками, благодаря которым рыба способна двигаться и удерживать равновесие. Лишенная плавников, она переворачивается брюшком вверх, поскольку центр тяжести помещается в спинной части.
Плавники — это кожистые выросты, опирающиеся на костные плавниковые лучи. Различают парные плавники, соответствующие конечностям высших позвоночных, и непарные (вертикальные).
Парные плавники — это грудные (p. pectoralis — Р) и брюшные (p. ventralis — V). Грудные плавники наиболее развиты у летающих рыб (позволяют им пролетать до 800 м) или ползающих по суше тропических Periophthalmys. Брюшные плавники сильно варьируют по положению: они могут смещаться вниз и вперед, под грудные (окунь) или даже на глотку (тресковые), превращаться в присоску (бычки, пинагор).
Грудные и брюшные плавники выполняют функцию стабилизаторов, несущих плоскостей, рулей, иногда органов движения. С их помощью рыба поддерживает тело в нужном положении: при удалении их рыба плавает наклонно (в сторону удаленных плавников), а при перерезке грудных — хвостом вверх.
Непарные плавники представлены спинным p. dorsalis — D, анальным p. analis — А и хвостовым p. caudalis — С.
Количество спинных плавников варьирует (у карпа — 1, у судака —2, у трески — 3). У скумбриевых за двумя спинными и анальным плавниками находятся многочисленные маленькие добавочные плавнички. У некоторых рыб может быть не один, а два анальных плавника. Так же варьируют функции плавников. У рыбы-прилипало спинной плавник превращен в присоску, а у рыбы-парусника, достигая высоты 1,5 м и возвышаясь над водой, служит парусом. Непарные плавники обеспечивают устойчивость тела.
Хвостовой плавник у подавляющего большинства рыб выполняет роль движителя, помогающего им плавать. Некоторые рыбы этого плавника не имеют (скаты).
Форма хвостового плавника также связана с образом жизни рыбы.
Неравнолопастный (гетероцеркальный) хвост большей частью сочетается с нижним ртом (акулы, осетровые).
У летающих рыб сильнее развивается нижняя часть хвоста, и это помогает им выпрыгивать из воды. (У некоторых глубоководных рыб нижняя лопасть хвостового плавника больше длины тела)
Форма хвостового плавника связывается и со скоростью плавания: у наилучших пловцов — тунцов — хвост полулунный; медленнее плавает сельдь, имеющая вилообразный хвост; ещё медленнее — сазан, судак, лосось, обладающие выемчатым хвостом; у малоподвижного налима хвост округленный и т. д.
У некоторых рыб (лососевые, корюшковые, хариусовые, косатковые, кошачьи сомы) позади спинного есть жировой плавник (adiposa), не имеющий костной опоры.
В количестве и положении плавников возможны вариации. Парных плавников нет у круглоротых (миноги, миксины). Змеевидная форма тела обычно сопровождается потерей брюшных плавников (Anguillidae), а иногда и грудных (Muraenidae).
Плавание рыб
Плавание рыб происходит благодаря работе поперечнополосатой мускулатуры, которая приводит в движение движители. Рыбы передвигаются в толще воды, создавая упор, т.е. отталкиваясь от среды, обладающей некоторой плотностью, вязкостью и податливостью.
Форма тела и его частей обусловливают возможность передвижения рыб (способы, скорости и т.д.). Взаимодействие формы тела (обтекаемой, сплющенной или иной), размеров, формы и расположения плавников позволяет рыбе наилучшим образом использовать гидродинамические силы, возникающие при движении. У хороших пловцов (акулы, осетровые) подъемная сила увеличивается за счет выпуклой спинной и уплощенной брюшной поверхности тела, грудных плавников и рыла, направленных под углом к движению. Движения хвоста и хвостового плавника перемещают тело вперед и изменяют направление движения в горизонтальной и вертикальной плоскостях.
Различия в форме тела и образе жизни рыб привели к их различиям в способах движения. Основные из них таковы:
· Движение при помощи боковых колебательных изгибов всего тела. Скорость движения невелика. Таким способом передвигаются придонные рыбы, имеющие удлиненное тело (угри, вьюны).
· Движение при помощи частых боковых колебательных движений задней части тела. Передняя часть тела рассекает воду, задняя является движителем. Рыбы имеют компактное тело и мощный хвостовой стебель. К этой группе относится большинство рыб.
Скорость плавания рыбы зависит от скорости движения плавника. Скорость плавания находится в прямой зависимости от частоты гребков, которая может достигать 20 Гц, но обычно не превышает 5 Гц.
Движение грудных плавников у большинства рыб поочередное типа «кроль», но у некоторых видов, например у карпа,— одновременное типа «брасса». Скорость плавания при работе одних только плавников небольшая, расход энергии тоже, по-видимому, небольшой, так как в работу вовлекаются небольшие массы мускулатуры плечевого пояса. Коэффициент полезного действия мускулатуры при таком типе плавания довольно высок.
При плавниковом плавании туловищно-хвостовая мускулатура не бездействует, она в какой-то степени напряжена для поддержания обтекаемой «позы». В противном случае тело начинает пассивно колебаться в набегающем потоке, как водоросль в потоке, или как флаг на ветру (так называемый флаттер), и это вызывает торможение.
Наибольшие скорости достигаются рыбами при периодическом волнообразном изгибании всего тела (ундуляции). Движителем в этом случае служит почти вся поверхность тела, за исключением негибкой головы. Активно сокращается почти вся туловищно-хвостовая мускулатура. Упор при ундуляционном плавании создается искривлением тела и движением локомоторной волны от головы к хвосту. Максимальная частота ударов хвоста при плавании некоторых видов достигает 30 Гц.
Данные разных авторов, приведенные ниже (Таблица 2), показывают, что наибольшие частоты плавательных движений доступны среди взрослых рыб хищникам, таким, как тунцы, и пелагическим: рыбам, таким, как атерина. Личинки рыб тоже имеют очень высокие частоты плавательных движений, чтобы выйти из области низких значений Re, где кинематическая вязкость воды очень высока и эффективность плавания имеет низкие значения.
Максимальные частоты ундуляции (плавательных движений) некоторых видов рыб
| Вид рыбы | Длина, см | Максимальная известная частота, Гц. | Вид рыбы | Длина, см. | Максимальная известная частота, Гц. |
| Тунец | 100 | 30 | Хамса | 12 | 30 |
| Вьюнок | 20 | 10 | Атерина | 7 | 30 |
| Бычок | 15 | 20 | Колюшка | 7 | 15 |
| Карп | 25 | 10 | Гольян | 7 | 15 |
Максимальные частоты ундуляции и соответствующие им максимальные, так называемые спринтерские, скорости могут поддерживаться в течение нескольких секунд. Далее наступает утомление и скорость резко падает. Поскольку максимальная скорость ундуляции у разных видов колеблется от 10 до 30 Гц, то максимальная скорость может колебаться от 7,5 до 22,5 длины тела в секунду.
У крупных хищников и мигрантов абсолютные значения максимальной скорости могут достигать больших величин. Двухметровая рыба, работающая с частотой 30 Гц и движущаяся со скоростью около 20 длин тела в секунду, развивает скорость 40 м/с, или 144 км/ч.
Существует некоторая неутомляющая частота мышечных сокращений, при которой времени между двигательными актами достаточно для полного восстановления работоспособности. При таких режимах движения происходят длительные миграции рыб. Неутомляющая средняя скорость в 5–10 раз меньше максимальной спринтерской скорости плавания.
Проведенные эксперименты и наблюдения в естественных условиях показали, что лососи не утомляются при скорости плавания 1–2 длины тела в секунду, т.е. до 5 км/ч. Тунцы — рекордсмены среди рыб; по скоростной выносливости могут длительное время двигаться со скоростью 3–4 длины тела в секунду, т.е. порядка 20 км/ч.
Стайерская скорость может поддерживаться несколько часов, но она все-таки не является неутомляющей, или крейсерской, скоростью. Наилучшие среди рыб пловцы — тунцы, ставриды, лососи — плавают в 2–3 раза быстрее и они выносливее, чем посредственные пловцы, такие, как осетровые, камбалы, бычки, угри.
Мускулатура рыб
Мышечную систему рыб, как и других позвоночных, разделяют мышечную систему тела (соматическую) и внутренних органов (висцеральную). Группы мышц головы управляют движениями челюстного и жаберного аппаратов (висцеральная мускулатура).
Мускулатура — это специализированная сократительная ткань, которая подразделяется на два основных типа: поперечнополосатую произвольную и гладкую непроизвольную мускулатуру.
Гладкая непроизвольная мускулатура.
Она работает независимо от волевых актов, хотя тоже иннервирована и подчиняется действию нервных и гормональных стимулов. Гладкая мускулатура способна к спонтанным ритмическим сокращениям даже вне организма. Например, полный цикл периодического движения желудка и кишечника рыб при комнатной температуре занимает несколько минут.

Сокращение гладкой мускулатуры стимулируется многократными нервными и электрическими импульсами. Быстрое растяжение гладких мышечных волокон под действием внешних сил, например растяжение желудка проглоченной пищей, вызывает ответное сокращение. В стенках полых органов (желудка, кишечника, пузырей) имеются продольные и кольцевые гладкие мышечные волокна. Гладкая мускулатура имеется также в селезенке, радужине глаза и в аппарате движения хрусталика глаза, в стенках кровеносных сосудов и жабрах.
Поперечнополосатая мускулатура
В мышечной системе тела выделяют мускулы туловища, головы и плавников. Внутренние органы имеют свою мускулатуру.
Мышечная система взаимосвязана со скелетом (опора при сокращении), с нервной системой (к каждому мышечному волокну подходит нервное волокно, и каждая мышца иннервируется определённым нервом). Нервы, кровеносные и лимфатические сосуды располагаются в соединительнотканной прослойке мышцы. Сама соединительнотканная прослойка в мышцах рыб в отличие от мышц млекопитающих невелика.
У рыб, как и других позвоночных, сильнее всего развита туловищная мускулатура. У настоящих рыб она представлена двумя большими тяжами, расположенными вдоль тела от головы до хвоста (большая боковая мышца — m. lateralis magnus) (рис. 12).
Поперечнополосатая произвольная мускулатура, или туловищно-хвостовая, является основным двигателем при плавании рыб. Деятельность ее в значительной мере определяется волевыми актами, хотя при плавании большую роль играет автоматизм.
Туловищно-хвостовая мускулатура рыб состоит из отдельных сегментов (миомеров), число которых равно числу позвонков. У рыб вытянутой формы число миомеров около 100 и больше (угорь), у рыб обыкновенной формы (карп, окунь, сельдь, лосось, камбала) примерно 50, у коротких рыб (бычки, спинороги, ставрида) около 25 миомеров.

Мускулатура костистой рыбы (окуня) (по Кузнецову, Чернову, 1972):
1 — миомеры, 2 — миосепты
Сегментарное строение характерно как для красной, или темной, латеральной мускулатуры, так и для белой, или светлой, составляющей большую часть всей мышечной массы. Несмотря на сегментарное строение, красная мускулатура выступает как единое целое. Все ее мышечные волокна направлены вдоль оси тела. Сокращение этих волокон вызывает латеральные изгибы тела. Белая мускулатура может быть разделена на: спинную — выше средней линии тела, и боковую — ниже средней линии тела.
Соединительнотканные элементы, отделяющие спинную мускулатуру от боковой и один миомер от другого, называются миосептами. К ним присоединяются мышечные волокна, с помощью которых сокращение мускулатуры передается костному скелету и вызывает локомоторные волнообразные движения рыбы.
Поперечно-полосатые волокна, а следовательно, и мышцы делят на красные и белые, различающиеся, как следует из названия, цветом. Цвет обусловлен наличием миоглобина — белка, легко связывающего кислород; миоглобин обеспечивает дыхательное фосфорилирование, сопровождающееся выделением большого количества энергии.

Светлая и темная мускулатура рыб во многом различна. Судя по данным электрофизиологии, при спокойном плавании рыбы в крейсерском режиме число сокращений волокон темной мускулатуры равно числу плавательных движений. Сокращения волокон белой мускулатуры носят нерегулярный характер.
Волокна красной мышцы (m. lateralis superficialis) узкие, тонкие, интенсивно кровоснабжаемые, расположенные более поверхностно (вдоль тела от головы до хвоста), содержат в саркоплазме больше миоглобина; в них обнаружены скопления жира и гликогена. Возбудимость их меньше, отдельные сокращения длятся дольше, но протекают медленней; окислительный, фосфорный и углеводный обмен интенсивнее, чем в белых.
К широких, более толстых, светлых — белых волокнах m. lateralis magnins миоглобина мало; меньше в них гликогена и дыхательных ферментов. Углеводный обмен происходит преимущественно анаэробно, и количество выделяемой энергии меньше. Отдельные сокращения сравнительно быстры. Мышцы быстрее сокращаются и утомляются, чем красные. Лежат они более глубоко. Красные мышцы постоянно деятельны. Они обеспечивают длительную или непрерывную работу органов. Именно они поддерживают постоянное движение грудных плавников, обеспечивают изгибы тела при плавании и поворотах, непрерывную работу сердца.
Они как бы поддерживают постоянную ритмичную работу темных мышц энергичными мощными движениями. Эти движения рыбы производят с определенными интервалами. Как правило, в течение секунды они движутся энергично, затем приблизительно столько же движутся по инерции или стоят.
Количество темных мышц у рыб различно. Особенно много их у постоянно плавающих пелагических рыб — тунцов, макрелей, ставрид, сельдей. У тунцов темная мускулатура расположена не только в виде тонкого подкожного слоя, как у большинства видов, но и в толще светлых мышц до самого позвоночника. Доля темной мускулатуры может достигать 20% всей мышечной массы.
Совсем мало темных мышечных волокон у рыб, ведущих малоподвижный образ жизни, таких, как скорпены, а также у рыб, передвигающихся в основном с помощью плавников.
Темные мышцы называются медленными, или тоническими, так как мышцы этого типа поддерживают у наземных позвоночных позу и общий тонус мышечной массы. Они иннервируются нервными волокнами малого диаметра, концевые контакты которых распределены по нескольку на каждое мышечное волокно, что позволяет более точно регулировать силу и скорость сокращения тонической мускулатуры.
Светлые мышцы еще называют фазическими, или быстрыми. Их мощность и скорость сокращения вдвое больше, чем у темных мышц. Для быстрых мышечных волокон пластинчатожаберных, двоякодышащих, хрящевых ганоидов и примитивных костистых рыб характерна иннервация каждого волокна одним нервным окончанием, а для большинства костистых рыб характерна множественная иннервация каждого волокна, так же как для волокон темной мускулатуры.
Особенность пищи и окружающей воды могут сильно изменить пищевую ценность рыбы: в заболоченных, тинистых или загрязненных нефтепродуктами водоемах рыбы имеют мясо с неприятным запахом.
Качество мяса зависит от диаметра мышечного волокна, количества жира в мышцах и других факторов. В значительной мере оно определяется соотношением массы мышечной и соединительной ткани, по которому можно судить о содержании в мышцах полно ценных мышечных белков (по сравнению с неполноценными белками соединительнотканной прослойки).
Это соотношение изменяется в связи с физиологическими особенностями организма и факторами внешней среды, в том числе с возрастом и условиями выращивания рыбы.
Электрические органы рыб
Электрические явления имеют собственное значение или сопровождают многие жизненные процессы, если происходит перемещение ионов и имеется несимметричное распределение ионов по разные стороны биологических мембран. Этообусловливает наличие электрических потенциалов и возникновение электрических токов.
Биотоки регистрируются при плавании рыб, дыхательных движениях. Электрокардиограммы рыб, несмотря на отличие сердца рыб от человеческого, имеют те же характерные «зубцы». Записи биотоков сетчатки глаза (ретины), называются ретинограммами. Запись биотоков головного мозга называется энцефалограммой.
Биотоки возникают при работе органов и тканей у разных животных, но только у рыб имеются специализированные органы, генерирующие электрический ток. Электрические токи, генерируемые рыбами, позволяют им общаться друг с другом, а также используются для поиска, локации, защиты и нападения.
Своеобразно измененными мышцами являются электрические органы. Эти органы у большинства имеющих их рыб развиваются из зачатков поперечно-полосатой мускулатуры. Они расположены по бокам тела и у современных рыб состоят из множества мышечных пластинок (у электрического угря их около 6000), преобразованных в электрические пластинки, переслаиваемые студенистой соединительной тканью. Нижняя часть пластинки заряжена отрицательно, верхняя — положительно. Разряды происходят под действием импульсов продолговатого мозга.
Электрические органы могут располагаться в разных частях тела, например у ската морской лисицы — на хвосте, у электрического сома — на боках.
Генерируя электрический ток и воспринимая силовые линии, искаженные встречающимися на пути предметами, рыбы ориентируются в потоке, обнаруживают препятствия или добычу с расстояния в несколько метров даже в мутной воде. Органы зрения в этих условиях помочь им не могут и редуцируются.
В соответствии со способностью к генерации электрических полей рыб разделяют на три группы:
1. Сильно электрические виды — имеют большие электрические органы, генерирующие разряды от 20 до 600 В. Основное назначение разрядов — нападение и оборона (электрический угорь, электрический скат, электрический сом).
2. Слабоэлектрические виды — имеют небольшие электрические органы, генерирующие разряды напряжением менее 17 В. Основное назначение разрядов — локация, сигнализация, ориентация (обитающие в мутных реках Африки многие мормириды, гимнотиды, некоторые скаты, нильская щука и нильский длиннорыл).
3. Неэлектрические виды — не имеют специализированных органов, но обладают электрической активностью. Генерируемые ими разряды распространяются на 10–15 м в морской воде и до 2 м в пресной. Основное назначение генерируемого электричества — локация, ориентация, сигнализация (многие морские и пресноводные рыбы: ставрида, атерина, окунь и др.).
ТЕМА 2: Нервная система
Нервная система объединяет и согласует деятельность всех систем организма, в результате чего организм способен правильно реагировать на изменения внешней и внутренней среды.
Нервная система состоит из центральной и периферической. В центральной нервной системе — происходит переработка поступающей информации и принимаются целесообразные решения, а периферическая нервная система передает информацию и точно адресованные команды.

Нервная система рыб представлена центральной нервной системой и связанной с ней периферической и вегетативной (симпатической) нервной системой. Центральная нервная система состоит из головного и спинного мозга. К периферической нервной системе относятся нервы, отходящие от головного и спинного мозга к органам. Вегетативная нервная система в основе имеет многочисленные ганглии и нервы, иннервирующие, мышцы внутренних органов и кровеносных сосудов сердца. Нервная система рыб по сравнению с нервной системой высших позвоночных характеризуется рядом примитивных черт.
Строение и функции нерва
Нерв представляет собой пучок нервных волокон — проводящих отростков нервных клеток, покрытых общей соединительнотканной оболочкой. Тела нервных клеток с ядрами и прочим клеточным содержимым находятся в центральной нервной системе и периферических ганглиях.
Нервная клетка — нейрон — представляет собой структурную единицу нервной ткани. От нейрона отходит один длинный отросток — аксон— и несколько более коротких ветвящихся отростков — дендритов. Аксон или совсем не ветвится, или ветвится при вхождении в иннервируемый орган.
Нервные волокна бывают безмякотные и мякотные, т.е. обладающие толстой изолирующей миелиновой оболочкой, образованной особыми клетками.
В безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны. В миелиновых нервных волокнах возбуждение распространяется скачкообразно от одного перехвата Ранвье к другому, как бы перепрыгивая через участки, покрытые миелиновой оболочкой. Эти расстояния достигают 1 мм. Скорость распространения нервного импульса по мякотным волокнам больше, чем по безмякотным, где она находится в прямой связи с толщиной волокна.
Повреждение нервного волокна нарушает проведение импульса. Возбуждение, передающееся по какому-либо нервному волокну, не передается на соседние нервные волокна этого же нервного ствола.
Окончания аксона теряют миелиновую оболочку и образуют своеобразные утолщения — синаптические бляшки.
В синапсах центральной нервной системы происходят контакты между аксоном одного нейрона и клеточным телом другого — аксосоматические синапсы; между аксоном одного нейрона и дендритом другого — аксодендрические синапсы; между аксоном одного нейрона и аксоном другого — аксоаксонные синапсы.
Периферические синапсы осуществляют контакт между аксоном двигательного нейрона и работающим органом. На одних клетках оканчиваются сотни или тысячи пресинаптических волокон, другие могут иметь один синапс. Синапсы могут быть возбуждающими или тормозящими в зависимости от того, активизируют они или подавляют деятельность органа.
|
|
|
