 |


Факторы, влияющие на интенсивность энергетических трат
|
|
|
|
Величина энергетических трат зависит от ряда причин внутреннего характера и от многих факторов внешней среды. Скорость обмена веществ зависит от размеров тела, подвижности, степени нервного возбуждения, накормленности.
На интенсивность обмена веществ сильное влияние оказывают температура, кислород, соленость воды, сезон, освещенность и динамика изменения светового дня. Влияние различных факторов на траты энергии обычно устанавливается на фоне величины так называемого стандартного обмена, т.е. в отсутствие питания, движения и нервного возбуждения.
Обеспечить условия полной неподвижности при определении обмена веществ у рыб удается далеко не всегда. Как правило, получить данные о тратах энергии у рыбы удается при некотором (обычном) уровне двигательной активности. Измеренная величина называется обычным, или рутинным, обменом. Рутинный обмен выше стандартного в 1,2–1,5 раза.
Траты энергии и вещества во время эмбрионального периода тоже не могут быть классифицированы как стандартный или основной обмен, так как эмбрион постоянно и интенсивно питается за счет запасов желтка.
За время эмбрионального периода расходуется некоторое количество вещества, составляющего яйцо. В крупных яйцах лососевых и осетровых рыб до выклева расходуется несколько процентов начальной массы вещества. У мелких яиц сельди, карповых, окуневых этот расход более заметен и достигает 25% сухой массы.
От оплодотворения до перехода на внешнее питание тратится приблизительно половина вещества яйца, а остальное превращается в тело эмбриона (Рис.).
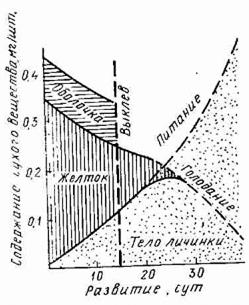
Рис. Баланс сухого вещества в течение эмбрионального и личиночного периодов развития сельди
|
|
|
Интенсивность обменных процессов зависит от величины организма. Самая высокая скорость энерготрат, потребления кислорода, расходования органических и минеральных веществ наблюдается у рыб на ранних стадиях развития. Величина рутинного обмена личинок карпа достигает 80 ккал/кг в сутки, что эквивалентно 17% всего вещества, составляющего тело. Очевидно, что личинки не в состоянии долго оставаться без корма. Максимальная величина рутинного обмена у радужной форели наблюдается при массе тела около 1 г (Рис.).
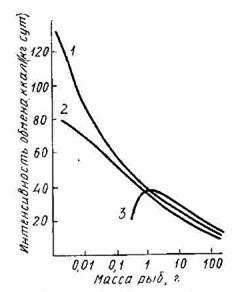
Рис. Величина рутинного (обычного) обмена рыб в зависимости от массы тела:
1 — по формуле Г.Г. Винберга; 2 — фактические данные по карпу; 3 — фактические данные по форели
При больших и меньших размерах тела уровень обмена у форели ниже. Следует заметить, что такой максимум обнаружен у многих животных — земноводных, млекопитающих. У человека максимальная интенсивность энерготрат наблюдается при массе тела порядка 10 кг. При массе тела рыб больше 1 г величина рутинного обмена монотонно понижается. Эту зависимость Г.Г. Винберг предложил выражать в виде степенной функции массы тела.
Для большинства обычных рыб умеренного климатического пояса (карповых, окуневых, лососевых, осетровых) при 20°С эта зависимость может быть выражена формулой:
Q=34W—0,2
где: W—масса тела рыбы, г; Q — величина рутинного обмена, ккал/(кг сут.).
Из этой формулы следует, что при 20°С рыбы с массой тела 1 г при обычной активности в отсутствие питания тратят в сутки 34 тыс. калорий энергии на 1 кг массы.
Интенсивность обмена веществ рыб, обитающих в других климатических зонах, в иных температурных условиях может иметь сходную величину в результате адаптационных процессов на биохимическом уровне (Рис). Однако общая тенденция такова, что в привычных условиях обитания тепловодные рыбы тратят больше энергии, чем холодноводные.

Рис. 4. Интенсивность обмена веществ (стандартный обмен) у рыб климатических зон (масса рыб 50–100 г): 1 — антарктические рыбы. 2 — полярные тресковые, 3 — полярные бычки и бельдюги, 4 — рыбы умеренного климата, 5 — тропические рыбы
|
|
|
Для каждого вида рыб имеется своя температурная зона обитания. Карп живет при температуре от 0 до 36°С, форель 0– 27°С. С повышением температуры до некоторого предела энерготраты увеличиваются. Температурная зависимость для сеголетков карпа выражается следующим рядом чисел (за единицу принят обмен при 20°С):
| Температура, °С | Величина энерготрат |
| 0 | 0,17 |
| 5 | 0,27 |
| 10 | 0,42 |
| 15 | 0,67 |
| 20 | 1,00 |
| 25 | 1,26 |
| 30 | 1,42 |
Температура является мощным фактором, влияющим на интенсивность жизнедеятельности рыб. Рыбы — водные животные, температура тела подавляющего большинства рыб соответствует температуре окружающей среды.
Одни виды обитают в приполярных водоемах, и даже 10°С — для них слишком высокая температура, при этом обменные процессы подвергаются угнетению. Другие рыбы живут в тропиках, где обычной температурой является 25–35°С. Температурное угнетение обмена веществ тропических рыб находится в области выше 40° С.
Главные объекты нашего рыбоводства — радужная форель и карп — различаются точкой температурного угнетения. Форель — объект холодноводного рыбного хозяйства. При температуре выше 20°С она начинает плохо себя чувствовать и интенсивность обмена у нее понижается. Обменные процессы карпа продолжают расти до 30°С.
Активное плавание вызывает увеличение расхода энергии и вещества. Влияние скорости плавания на расход энергии выражается квадратической зависимостью. Приблизительную энергетическую «стоимость» плавания рыб можно выразить формулой [ккал/(кг-ч)]:
Qплав=10v2/L
где: v — скорость плавания, см/с; L — длина рыбы, см.
В спокойном состоянии рыбы тратят энергии 0,2– 1 ккал/(кг-ч), а при крейсерской (стайерской) скорости плавания расход энергии возрастает до 2–7 ккал/(кг-ч) (Таблица).
Таблица. Возрастание трат энергии у рыб при повышении двигательной активности по сравнению с покоем
| Рыба | Экологическая ситуация | Траты энергии ккал/(кг-ч) | |
| в покое | при движении | ||
| Лососи | Нерестовая миграция | 0,20 | 0,02–2,0 |
| Осетровые | Нерестовая миграция | 0/24 | 0,025–1,0 |
| Угорь | Нерестовая миграция | 0,20 | 0,04–0,26 |
| Сельдь | Миграция | 0,25 | До 2,4 |
| Судак | Активная охота | 0,30 | 0,12 |
| Ставрида | Активная охота | 0,80 | 0,13 0,5 |
| Тунец | Дальняя миграция | 0,50 | |
| Тунец | Активность | 5,0 | |
| Атерина | Питающаяся стая | 7,0 | |
|
|
|
При бросках расчетные траты энергии возрастают в раде случаев в десятки раз (Таблица) и могут достигать величин, эквивалентных 250 ккал/(кг-ч). Естественно, что такие расходы энергии могут происходить только в течение нескольких секунд.
Таблица. Траты энергии у рыб при максимальных скоростях
| Рыба | Длина тела, см | Скорость, см/с | Траты, ккал/(кг ч) |
| Карп | 25 | 197 | 14 |
| Вьюн | 15 | 150 | 16 |
| Хамса | 12 | 80 | 25 |
| Форель | 30 | 750 | 80 |
| Ставрида | 12 | 300 | 250 |
Суммарное выражение прироста трат вещества (усиление распада тканевых веществ и метаболизирование части вещества пищевого происхождения) во время усвоения пищи называется специфическим динамическим действием пищи (СДД). Это действие внешне выражается в усилении потребления кислорода и увеличении выделения продуктов обмена.
Потребление кислорода, выделение углекислоты и аммиака начинают увеличиваться почти сразу после приема пищи и через несколько часов достигают максимума, а затем постепенно снова понижаются до уровня рутинного обмена. Максимальная интенсивность дыхания после приема пищи может в несколько раз превышать обычный уровень.
Если рыба питается постоянно и постоянно происходит усвоение питательных веществ, то уровень ее обмена повышен по сравнению с «голодным уровнем».
Питание увеличивает интенсивность катаболических реакций организма рыбы. Среднесуточная интенсивность потребления кислорода и выделения продуктов обмена возрастает пропорционально величине суточного рациона. Это увеличение интенсивность трат вещества и энергии в результате обменных процессов составляет 15–20% общей энергии суточного рациона, или до 1/3 энергии усвояемой части рациона.
Опыты с использованием радиоактивного углерода показали, что скорость обмена накопленных в теле веществ возрастает в результате СДД довольно слабо — не более чем в 1,2 раза. Основная часть прироста выделяемых продуктов метаболизма зависит от пищи, находящейся в данный момент в кишечнике. Их энергия тратится на превращение веществ пищи в вещества тела.
|
|
|
Состав расходуемых веществ зависит от химического состава тела рыбы. У рыбы высокой упитанности в отсутствие внешнего питания энергетические траты на 60–70% покрываются за счет жира и на 30–40% —за счет белка. По мере истощения запасов жира, например во время зимовки или вынужденного голодания, увеличиваются траты белка и уменьшаются траты жира (Рис).

Рис. 5. Величина расхода белка по мере истощения жировых запасов при голодании мальков карпа массой 1 г при 30°С: 1 — белок; 2 — жир
На происхождение субстратов биологического окисления при питании большое влияние, очевидно, оказывает состав корма. Однако особенностями рыб по сравнению с другими животными является требование высокого содержания белка в усвояемой части корма и особая важность аминокислот как субстрата окисления.
Если птиц и жвачных животных можно кормить пищей, содержащей 12% белка, то форели и карпу требуются корма, в усвояемой части которых содержится 30–40% белка.
Углеводы составляют не более 1% массы тела рыб. В соответствии с этим невелико среднее значение их доли в покрытии энергетических трат основного обмена. Однако роль углеводов в производстве энергии может временами сильно возрастать.
Если рыба накормлена пищей, богатой хорошо усвояемыми углеводами, то во время их усвоения будет происходить нагрузка внутренней среды сахарами и доля их в энергетическом катаболизме повысится. Углеводы, а именно глюкоза, могут служить промежуточным продуктом при производстве энергии из других химических соединений — различных аминокислот и жирных кислот.
В организме рыбы глюкоза довольно легко превращается в гликоген, хотя у рыб связывание глюкозы идет гораздо медленнее, чем у млекопитающих. Гликоген запасается не только в специализированном депо — печени, но и прямо в тех тканях, где непосредственно используется глюкоза,— в мышцах, почках, ооцитах, мозгу.
Гликоген легко подвергается ферментативному гидролизу, быстро снабжая клетки тела большим количеством одинаковых, легко используемых молекул глюкозы, которые используются до того, как начнется более инертный процесс мобилизации жировых веществ. Существует даже такое образное выражение — «жиры сгорают в пламени углеводов».
Короткие неожиданные всплески мышечной активности осуществляются за счет глюкозы, которая способна давать энергию за счет анаэробного (бескислородного) распада до молочной кислоты, хотя при этом тратится в 20 раз больше вещества, чем при окислении.
|
|
|
Роль глюкозы особенно велика при работе белой туловищной мускулатуры, а также при перенесении временного недостатка кислорода.
|
|
|
