 |


Жабры как орган осморегуляции и экскреции
|
|
|
|
В жаберном эпителии миног, акул и костистых рыб имеются особые клетки, очень богатые митохондриями т.е. способные к энергоемким процессам. Это так называемые хлоридные клетки.
У пресноводных рыб и у эвригалинных и проходных рыб в пресной воде хлоридные клетки извлекают натрий из пресной воды, в которой он всегда содержится хотя бы в следовых количествах. Через жабры рыб проходит в сутки порядка 1 м3 воды, где содержится до 100 г хлоридов. В соленой воде хлоридные клетки выводят лишний натрий из организма.
Перестройка работы клеток требует некоторого времени. Угри, адаптированные к пресной воде, начинают усиленно секретировать натрий через 1–2 ч после помещения их в соленую воду. При работе на полную мощность хлоридные клетки угря способны вывести за час 1/3 натрия, содержащегося в организме.
Разные виды отличаются различной адаптационной способностью. У карпа, карася, морского окуня, тунца эта способность ограниченна — они довольно стеногалинны. У лососевых эта способность развивается в онтогенезе, они становятся способными перейти в морскую воду по достижении так называемой покатной стадии.
Жабры являются основным органом экскреции из организма аммиака, но не мочевины. Например, у бычка-рогатки во время прохождения крови через жабры содержание аммиака уменьшается втрое, а содержание мочевины заметно не изменяется. Поэтому жабры являются главным местом экскретирования продуктов катаболизма белка у аммонотелических рыб (главным метаболитом белка у которых — является аммиак): миног, миксин, пресноводных и костистых морских рыб.
Ректальная железа акуловых рыб
В заднем отделе кишечника акулы имеется специфический орган, функцией которого является экскреция лишних хлоридов. Он развивается как выпячивание задней кишки и располагается в полости тела над кишечником. Это так называемая ректальная железа. Она обильно снабжается кровеносными сосудами и имеет в своем составе много богатых митохондриями и микротрубочками клеток. Выделения ректальной железы акул содержат вдвое больше натрия и хлора, чем кровь, и почти совсем не содержат мочевины. Ректальная железа, таким образом, является специализированным органом экскреции хлоридов.
|
|
|
Роль пищеварительного тракта в осморегуляции
Пищеварительный тракт играет заметную роль в балансе воды и солей у рыб. Особенно большое значение кишечник имеет для морских костистых и миног. Они, как известно, пьют морскую воду для компенсации осмотических потерь влаги.
Большинство морских рыб выпивает от 80 до 360 мл морской воды на 1 кг массы тела в сутки. Потребление некоторых рыб превышает их собственную массу (тиляпия, некоторые виды бленниид). При этом 60–80% этой воды всасывается в кишечнике.
Вместе с водой всасываются почти полностью одновалентные ионы (натрий, калий, хлор) выпитой морской воды. В кишечнике остаются в свободном состоянии ионы магния и сульфата, а также нерастворимые карбонаты кальция и магния, которые задерживаются мукоидной выстилкой кишечной стенки.
Через пищеварительный тракт морских и пресноводных рыб экскретируется значительное количество соединений кальция, магния и фосфора пищевого и эндогенного происхождения
Усвоение воды в кишечнике морской рыбы происходит быстрее, чем у пресноводной рыбы, несмотря на то, что у морской рыбы это всасывание происходит против осмотического градиента, создаваемого разностью концентраций между проглоченной морской водой и кровью. Это доказывает, что кишечное всасывание воды у морских рыб есть процесс активный, требующий затрат энергии.
|
|
|
Пресноводные рыбы теряют некоторое количество воды в процессе пищеварения, так как химус и экскременты, как правило, содержат больше влаги, чем пища, особенно сухие гранулированные корма.
ТЕМА 9: Воспроизводительная система рыб
Функцией гонад рыб является продуцирование яиц (икринок) и сперматозоидов. Икринки и сперматозоиды образуются из первичных половых клеток, обособляющихся еще во время эмбрионального периода онтогенеза и имеющих размер 9–20 мкм. Длина зрелых сперматозоидов достигает 30–60 мкм, яйца же рыбообразных имеют самые различные размеры — от долей миллиметра до нескольких сантиметров (яйцекладущие скаты).
Пол у рыб
Часто система определения пола различна у близких видов, например у разных видов угрей и даже у разных популяций одного вида (пецилии). Внешние признаки женского или мужского типа развития выражаются в первую очередь в развитии гонад — половых клеток и структурных тканей. При внешнем осмотре рыбы и ее внутренних органов в раннем периоде определить пол рыбы, как правило, не представляется возможным и особи классифицируются как ювенильные.
На самом деле, ранние стадии развитая гонад довольно сложны и многообразны. Первичные половые клетки имеют способность развиваться, как по мужскому, так и по женскому пути.
Некоторым рыбам свойствен ювенильный гермафродитизм — у них развиваются как мужские, так и женские клетки, одни из которых впоследствии отмирают. Широко распространены явления протоандрии и прототинии, когда на определенной стадии развития все половые клетки развиваются по мужскому или по женскому пути.
Соотношение женских и мужских половых гормонов в организме рыбы обусловливает путь развития первичных половых клеток. И при женском, и при мужском типе развития они проходят сложную схему многократных делений и роста. Яйцеклетки растут в окружении системы питательных клеток, образующих вокруг них фолликулы.
Питательные клетки находятся и среди растущих сперматозоидов. Интерстициальные клетки гонад кроме питательной и опорной функций имеют еще эндокринную функцию. Гонады нередко называют еще половыми железами, их секреты оказывают мощное влияние не только на половые клетки и фолликулы, но и на весь организм.
|
|
|
У рыб известно несколько типов функционального гермафродитизма. Особенно много примеров этого явления среди окунеобразных, светящихся анчоусов и карпозубых. Есть рыбы, которые в более раннем возрасте являются самками, а затем функционируют как самцы, а для некоторых видов характерна обратная картина. В соответствии с этим в гонадах рыб развиваются женские и мужские половые клетки, а особям свойственно поведение мужского или женского типа.
Среди морских окуней имеются виды, у которых в гонадах одновременно развиваются и яйцеклетки, и сперматозоиды, а тип репродуктивного поведения может меняться в течение нескольких минут. Вмсте с тем меняется и характерная брачная окраска.
Особенности строения тела и окраски, характерные для репродуктивного периода, обычно служат надежным критерием при рассортировке зрелых рыб по половому признаку. У лососей признаком самцов является более яркое выражение брачных изменений в строении тела (горбатость), челюстей (крючковатостъ, зубатость), окраски кожи. Самцы вьюна легко различаются на ощупь по подкожным желвакам на боках тела в области хвостового стебля. У линя самцы и самки различаются развитием брюшных плавников. У самцов гольянов, плотвы и некоторых других карповых рыб на голове образуется специфическая жемчужная сыпь.
Пол многих видов рыб определяют по форме мочеполового отверстия. У живородящих пецилид половой диморфизм довольно рано проявляется в особом строении анального плавника самцов, который служит приспособлением для внутреннего оплодотворения. Такую же роль играют придатки брюшных плавников у акуловых.
Важным моментом принадлежности к тому или другому полу является тип репродуктивного и родительского поведения. В простейшем случае репродуктивное поведение сводится к образованию нерестовых скоплений и совместному вымету половых продуктов.
Более сложное поведение выражается в преследовании самцами самок (карп, щука). Еще более сложное поведение наблюдается в том случае, когда самцы подготавливают место для нереста и инкубации икры (гнездо) и приманивают самку.
|
|
|
Во время репродуктивного периода в поведении самцов часто появляются или обостряются элементы агрессивности, охраны территории, черты демонстративного поведения, ухаживания.
После вымета икры у многих видов наблюдается та или иная форма заботы о потомстве: обмахивание развивающейся икры, ношение ее во рту, ухаживание за личинками и мальками. Все эти формы поведения являются результатом включения врожденных стереотипов поведения под действием гормональной системы, одним из звеньев которой являются сами гонады.
Овогенез и сперматогенез
В процессе последних стадии развития в яйцеклетках накапливается желток и образуется оболочка, а у сперматозоидов образуется хвост, система движения и система внедрения в яйцеклетку. На стадиях дробления половых клеток гонады незначительны по величине, однако ко времени нереста и ястыки, и семенники увеличиваются в размерах (Рис.).

Рис. Изменение относительной массы гонад у балтийской трески в течение года
В это время накопление в них вещества становится существенным в общем балансе вещества и энергии в организме. Изменение относительного веса гонад у самцов и самок одного вида, как правило, происходит параллельно.
Несовпадение максимумов коэффициентов зрелости может наблюдаться у живородящих рыб с внутренним оплодотворением, так как осеменение может иметь место еще до полного созревания гонад у самок. В этом случае сперматозоиды сохраняются в яичниках самок до сроков оплодотворения.
Относительный вес зрелых гонад обычно больше у самок, чем у самцов. Однако у тресковых рыб семенники могут быть даже несколько больше, чем ястыки. Общая масса гонад у рыб различна. У некоторых видов масса гонад может достигать трети общей массы готовой к нересту рыбы (Таблица). Обычно в расцвете репродуктивного возраста рыба производит в год количество половых продуктов, составляющих около 30% ее массы.
Таблица. Масса зрелых гонад у некоторых рыб,%
| Вид рыбы | Самки | Самцы |
| Белый толстолобик | 10–15 | 0,1–0,5 |
| Карп | 10–25 | 2–9 |
| Радужная форель | 10–15 | 5–8 |
| Морской окунь | 15 | 0,6 |
| Нототения | 20 | 1 |
| Треска | До 30 | До 30 |
| Аю | 25 | 10 |
У многократно нерестующих тропических рыб годовое количество икры может превосходить массу тела. Значительная часть половых клеток резорбируется и служит материалом для построения выживающих клеток. Часть клеток в незрелых гонадах образует фолликулы и питающие клетки овоцитов и сперматозоидов. Питательные вещества поступают в половые железы по пронизывающим их кровеносным сосудам.
|
|
|
Гонады рыб обычно имеют вытянутую форму и подвешены в полости тела на брыжейках. При овуляции яйцеклетки выпадают из фолликулов в полость ястыка или прямо в полость тела и выводятся во внешнюю среду. Прямо в полость тела из оболочки гонад икра и сперма выливаются у круглоротых. Затем через поры попадают в мочевой синус или в мочеиспускающий канал и выводятся наружу через клоаку или урогенительную папиллу.
Гонодукт отсутствует у круглоротых. У осетровых и акуловых дня выведения икры или яиц служат мюллеровы протоки, сформированные из мочевых протоков головной почки. Сперма из них выводится по почечным каналам, которые у акул превращаются в лейдигов орган (Рис).

Рис. Строение яичников рыб: а — акула; б — лосось; в — сельдь: 1 — яичник; 2 — воронка яйцевода; 3 — почки, 4 — ректальная железа; 5 — кишка; 6 — мочеточники; 7 — клоака; 8– абдоминальные поры; 9 — матка; 10 — скорлупковая железа; 11– пузырь; 12 — анус; 13 — половое отверстие
У большинства костистых рыб каждый ястык имеет свой яйцевод, заканчивающийся генитальной порой в клоаке. У лососевых в связи с большой величиной икринок ястыки при созревании разрываются, и икра оказывается в полости тела, откуда она через воронку широкого яйцевода выводится во внешнюю среду.
Семенники костистых рыб имеют собственные семяпроводы, открывающиеся в мочеточник. При спермации, зрелые спермин освобождаются из ткани семенника и выводятся во внешнюю среду через семяпроводы.
Овуляция и спермация характеризуются определенными цитологическими процессами. С позиции физиологии эти процессы представляют собой функциональный метаморфоз ткани под действием гормонов. Фолликулярная ткань распадается, выдавливает из себя овоциты и спермии, образуется овариальная и спермальная жидкость.
Овуляция происходит или сразу во всем ястыке, и тогда рыба способна к одноразовому нересту, или она захватывает только часть созревающих овоцитов, и тогда рыба способна к многократному порционному нересту. Спермация происходит, как правило, в меньшем масштабе, зрелые спермин обнаруживаются в семенниках длительное время, и самцы обычно участвуют в нересте неоднократно.
Плодовитость самок рыб очень сильно различается в зависимости от величины икринок. Акуловые имеют очень невысокую плодовитость, но большая часть их потомства остается живой в результате развития внутри материнского организма или внутри прочной яйцевой оболочки. Желток яиц скатов и акуловых похож по форме и составу на птичий, он окружен настоящим «белком». Яйца акуловых развиваются очень долго — иногда до двух лет. Мелкая икра плодовитых рыб гибнет массами. Плодовитость костистых рыб колеблется в широчайших пределах. У обладающих крупной икрой видов — лососей, нототений — на каждый грамм массы приходится по одной икринке, у живородящих на 1 г тела приходится по нескольку десятков икринок, у карповых — по нескольку сотен, а у некоторых морских рыб, обладающих очень мелкой икрой, на 1 г тела приходятся тысячи икринок.
Количество спермы и содержание в ней спермиев различаются у разных рыб очень сильно. Общее количество спермы, выделяемой рыбой за репродуктивный период может превышать массу ее семенников, так как самцы могут участвовать в нересте многократно, и в это время ткань семенников продолжает продуцировать сперму.
У производителей радужной форели в течение 40 дней нерестового периода можно получить до 77 мл спермы (20 эякулятов),. у щуки гораздо меньше — до 6 мл (8 эякулятов), у крупных производителей толстолобика можно получить за один раз до 25 мл спермы, у осетровых — до 1 л.
Объем сперматозоидов в сперме (сперматокрит) тоже весьма различен. У лососевых сперматозоиды составляют 25% спермы, у карпа — 45%, у морского карася 77%, у некоторых камбал — даже 97%. В семенниках спермин неподвижны. Их активация происходит при разбавлении секретом придатка семенника — эпидимиальной жидкостью.
Для некоторых рыб необходимы особые условия активации. Например, спермин форели не активируются в кислой среде. Инактиваторами подвижности спермиев у некоторых видов являются ионы калия, которых много в спермальной жидкости.
Оплодотворение
Способность к оплодотворению после попадания в воду сохраняется у икринок ограниченное время. Это время обычно длится не более 2–3 мин, а часто и того меньше. Спермин рыб не обладают таксисом, т. е. способностью активно искать яйцеклетки, они находят их согласно статистическим закономерностям в результате своей многочисленности и подвижности. Двигательная активность спермиев в воде быстро возрастает до максимума (150 с), а Затем постепенно понижается (Рис.)
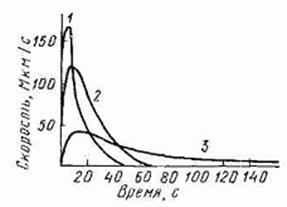
Рис. Динамика подвижности спермиев рыб 1 — осман, 2 — форель, 3 — чебак
За время движения спермин может самостоятельно продвинуться всего на 20–100 собственных длин, т.е. не более 1см. Проникновение спермия в яйцо происходит через микропиле — специальное воронкообразное отверстие во внешней оболочке.
Кроме микропиле во внешней оболочке костистых рыб имеется множество более мелких пор, пронизывающих ее насквозь. На разрезе эти поры создают впечатление исчерченности, поэтому оболочка икринки называется еще лучистой оболочкой. Сквозь поры лучистой оболочки легко проникают молекулы воды, но не проникают макромолекулы.
Перивитиллиновое пространство икринки образуется вне зависимости от оплодотворения. Вода проникает под оболочку и вызывает набухание кортикальных (корковых) альвеол, особых пузырьков, наполненных коллоидом. Альвеолы лопаются, коллоид, набухая, увеличивается в объеме и растягивает податливую в это время лучистую оболочку. Под ней образуется перивителлиновое пространство, защищающее зародыш от механических повреждений Образование перивителлинового пространства препятствует проникновению спермин и оплодотворению.
Сохранение икры и спермы рыб вне организма возможно в течение нескольких часов в сухой посуде и в помещении с низкой положительной температурой (Таблица). Более длительное сохранение спермы, например, для проведения скрещивания рыб, нерестящихся в разные календарные сроки, возможно при замораживании в специальной среде до температуры жидкого азота. Имеется опыт сохранения спермы даже в течение года.
Таблица. Сохранение спермы рыб
| Вид рыбы | Охлаяедение, °с | Длительность хранения при охлаждении, ч | Глубокое замораживание | |
| Длительность хранения, сут. | % оплодотворения | |||
| Сельдь | — | — | 180 | 85 |
| Треска | - | - | 1,5 | 89 |
| Кижуч | 8–10 | 8 | 7 | 70 |
| Радужная форель | 8–10 | 8 | 7 | 78 |
| Семга | 3–4 | 36–38 | 365 | 85 |
| Карп | -05 | 21–48 | 19 | 70 |
| Кефаль | 5 | 75 | 38 | 16 |
Зрелая овулировавшая икра может некоторое время сохранять способность к оплодотворению, если ее сохранять прямо в теле выловленной рыбы или в овариальной жидкости. Икра форелей и лососей может при нулевой температуре сохраняться: несколько суток, икра сиговых и карповых — несколько часов.
Многие акулы, скаты и некоторые костистые рыбы приспособились к внутреннему оплодотворению. Внутриутробное развитие может иметь различную степень совершенства. У сома трахихористеса сразу после внутреннего оплодотворения происходит вымет оплодотворенных икринок.
Внутреннее оплодотворение у морских окуней из рода себастодес также не имеет своей целью внутриутробное развитие. Сперма этих окуней сохраняется в полости тела до созревания яйцеклеток. По-видимому, внутреннее оплодотворение способствует в данном случае полноте оплодотворения икринок. Вслед за оплодотворением довольно быстро следует вымет икры во внешнюю среду, где происходит развитие
У морской бельдюги наблюдается настоящее живорождение. Желток у эмбрионов бельдюги очень маленький, и питательная среда для развития эмбрионов — эмбриотроф — формируется в полости гонад из погибших икринок и эмбрионов. У циматогастера из семейства эмбиотоцид внутриутробное развитие длится столь долго, что самцы рождаются половозрелыми.
Эмбриотроф образуется из остатков спермы, отмирающих овоцитов и эмбрионов. Всасывание питательных веществ происходит через поверхность желточного мешка, а дыхание осуществляется через сильно развитые лучи плавников Довольно хорошо развит процесс внутриутробного развития у живородящих рыб — пецилий, гуппи, молинезии и др. Эмбрионы этих рыб снабжены внешней кровеносной сетью для осуществления дыхания, а окружающие эмбрион фолликулы образуют питающую псевдоплаценту.
Особенно высокого совершенства достигает внутриутробное развитие у акул. Эмбрионы акуловых рыб рождаются вполне сформированными, их длина может достигать 20 см и даже больше у крупных видов. Развитие происходит в специальных матках, снабженных питающими образованиями наподобие плаценты млекопитающих,
|
|
|
